")
Abstract
Curcumin (diferuloylmethane) is a polyphenol derived from the plant Curcuma longa, commonly called turmeric. Extensive research over the last 50 years has indicated this polyphenol can both prevent and treat cancer. The anticancer potential of curcumin stems from its ability to suppress proliferation of a wide variety of tumor cells, down-regulate transcription factors NF-κ B, AP-1 and Egr-1; down-regulate the expression of COX2, LOX, NOS, MMP-9, uPA, TNF, chemokines, cell surface adhesion molecules and cyclin D1; down-regulate growth factor receptors (such as EGFR and HER2); and inhibit the activity of c-Jun N-terminal kinase, protein tyrosine kinases and protein serine/threonine kinases. In several systems, curcumin has been described as a potent antioxidant and anti-inflammatory agent. Evidence has also been presented to suggest that curcumin can suppress tumor initiation, promotion and metastasis. Pharmacologically, curcumin has been found to be safe. Human clinical trials indicated no dose-limiting toxicity when administered at doses up to 10 g/day. All of these studies suggest that curcumin has enormous potential in the prevention and therapy of cancer. The current review describes in detail the data supporting these studies.
Abbreviations
LDL, low density lipoprotein; HIV, human immuno-deficiency virus; 102, singlet oxygen; HVEC, human vascular endothelial cells; HVSMC, human vascular smooth muscle cells; PBMC, peripheral blood mononuclear cells; IL, interleukin; PDGF, platelet-derived growth factor; VSMC, vascular smooth muscle cells; PHA, phytohemagglutinin; PARP, polyadenosine ribose polymerase; ICAD, inhibitor of caspase-activated deoxyribonuclease; AIF, apoptosis-inducing factor; ROI, reactive oxygen intermediates; GSH, reduced form of gluthathione; PTK, protein tyrosine kinase; EGF, epidermal growth factor; EGFR, epidermal growth factor receptor; FPTase, farnesyl protein transferase; NF-κ B, nuclear factor-κ B; TNF, tumor necrosis factor; LT, lymphotoxin; ICAM-1, intracellular adhesion molecule-1; VCAM-1, vascular cell adhesion molecule-1; ELAM- 1, endothelial leukocyte adhesion molecule-1; COX2, cyclooxygenase-2; MMP, matrix metalloproteinase; ÈNOS, inducible nitric oxide oxidase; PMA, phorbol myristate acetate; AP-1, activating protein-1; πκ B•, inhibitory subunit of NF-κ B; UV, ultraviolet; JNK, c-jun N-terminal kinase; MAPK, mitogen-activated protein kinase; ERK, extracellular receptor kinase; IC50, inhibitory concentration (50%); MEKKI, mitogen-activated protein kinase kinase; TAKI, TGF-beta activated kinase-1; HPK1, human progenitor kinase 1; MAPKKK, mitogen-activated protein kinase kinase kinase; PKA, protein kinase A; PKC, protein kinase C; cPK, protamine kinase; PhK, phosphorylase kinase; AK, autophosphorylation-activated protein kinase; Ki, inhibition constant; TPA, tumor promoting agent; PKA, protein kinase A; cAK, autophosphorylation-activated protein kinase; CDPK, Ca2+-dependent protein kinase; LOX, lipoxygenase; HETE, hydroxyeicosatetraenoic acid; PGE, prostaglandin E, CD, chenodeoxycholate; CDK, cyclin-dependent kinase; SCC, squamous cell carcinoma; EC, endothelial cells; bFGF, basic fibroblast growth factor; VEGF, vascular endothelial growth factor; FGF, fibroblast growth factor; TGF-ß1, transforming growth factor-ß1; uPA, urokinase-activated plasminogen activator; TLC, total leukocyte count; GST, gluthathione S-transferase; HPLC, high pressure liquid chromatography; DMBA, 7, 12 dimethylbenz [a] anthracene; PhIP, 2-amino 1-methyl-6-phenylimidazo[4,5-b]pyridine; NNK, 4-(methyl-nitrosamino)-1-(3-pyridyl)-l-butanone; ODC, ornithine decarboxylase; PCNA, proliferating cell nuclear antigen; EBV, Epstein-Barr virus; B[a]P, benzo[a]pyrene; APC, antigen-presenting cells; BrdU, bromo-deoxyuridine; DHC, dihydrocurcumin; THC, tetrahydro-curcumin; HHC hexahydrocurcumin; i.v., intravenous, i.p., intraperitonial; i.g., intragastric.
Introduction
Curcumin, derived from turmeric (vernacular name: Haldi), is a rhizome of the plant Curcuma longa. The medicinal use of this plant has been documented in Ayurveda (the Indian system of medicine) for over 6000 years. It is commonly used as a spice, flavoring agent, food preservative, coloring agent, or for decoration. Extensive investigation over the last five decades has indicated that curcumin reduces blood cholesterol (1 -7), prevents LDL oxidation (8- 10), inhibits platelet aggregation (11, 12), suppresses thrombosis (13) and myocardial infarction (14-17), suppresses symptoms associated with type II diabetes (18-22), rheumatoid arthritis (23), multiple sclerosis (24) and Alzheimer (25, 26), inhibits HIV replication (27-31), enhances wound healing (32-34), protects from liver injury (35), increases bile secretion (36), protects from cataract formation (37), and protects from pulmonary toxicity and fibrosis (38-41), antileishmaniasis (42 – 44) and is an anti-atherosclerotic (45, 46). Additionally there is extensive literature that suggests that curcumin has potential in the prevention and treatment of cancer. The current review focuses on the studies that implicate curcumin in the prevention and therapy of cancer. We present several in vitro and in vivo preclinical and clinical results that suggest that curcumin has a great potential in the treatment of cancer. Certain aspects of curcumin have been reviewed (47-52), but in spite of numerous recent studies on its biology and mechanism of action, no comprehensive review has yet appeared on this subject.
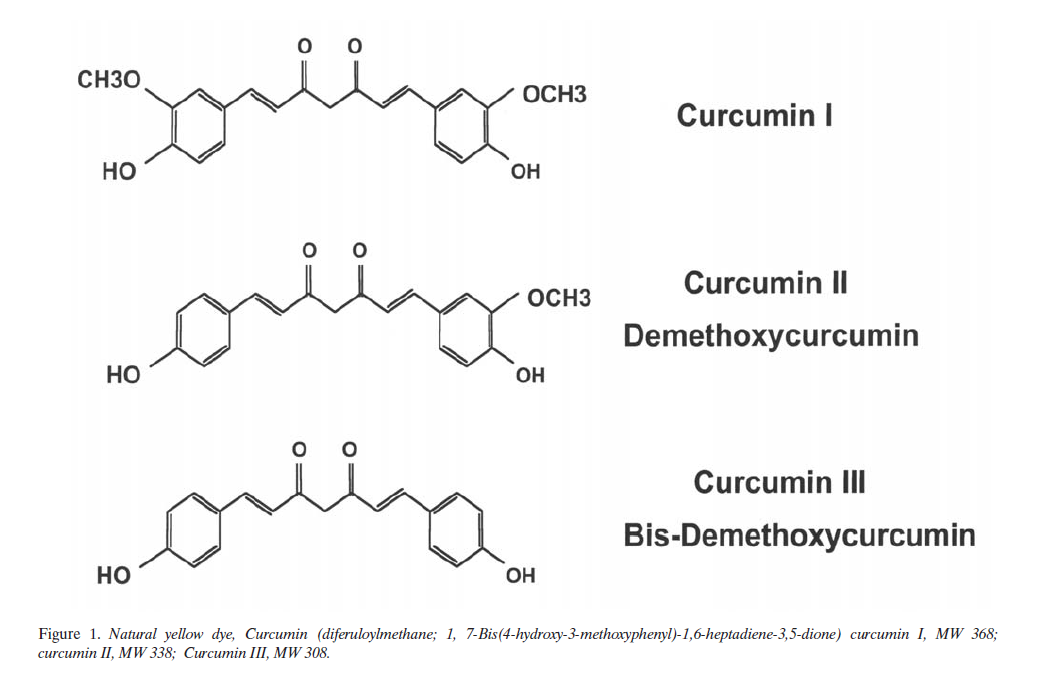
Chemistry of Curcumin
Curcumin (diferuloylmethane; see Figure 1) is a natural yellow orange dye derived from the rhizome of Curcuma longa Linn, an East Indian plant. It is insoluble in water and ether but is soluble in ethanol, dimethylsulfoxide and other organic solvents. It has a melting point of 183ÆC and a molecular weight of 368.37. Commercial curcumin contains three major components: curcumin (77%), demethoxy-curcumin (17%), and bisdemethoxycurcumin (3%), together referred to as curcuminoids (Figure 1). Spectro-photometrically, curcumin absorbs maximally at 415-420 nm in acetone and a 1% solution of pure curcumin has an optical density of 1650 absorbance units. It has a brilliant yellow hue at pH 2.5 to 7.0, and takes on a red hue at pH > 7.0. Curcumin fluorescence is a broad band in acetonitrile (Ïmax = 524 nm), ethanol (Ïmax = 549 nm), or micellar solution (Ïmax = 557 nm) (53). Curcumin produces singlet oxygen (1O2) upon irradiation (Ï > 400 nm) in toluene or acetonitrile (pHi = 0.11 for 50 ÌM curcumin); in acetonitrile curcumin also quenched 102 (kq = 7 x 106 M/S). 1O2 production was about 10 times lower in alcohols. Recently, Das and Das have studied the 1O2 quenching activity of curcumin in detail (54). Curcumin photogenerates superoxide in toluene and ethanol. In contrast, it quenches superoxide ions in acetonitrile (55).
Curcumin is also phototoxic to mammalian cells, as demonstrated in a rat basophilic leukemia cell model, and this phototoxicity likewise requires the presence of oxygen (56). The spectral and photochemical Aproperties of curcumin vary with environment, resulting in the potential for multiple or alternate pathways for the execution of photodynamic effects. For example, curcumin photogenerates singlet oxygen and reduced forms of molecular oxygen under several conditions relevant to cellular environments.

Tonnesen examined the kinetics of pH-dependent curcumin degradation in aqueous solution (57). A plot of the rate constant against pH indicated the pKa values of the acid protons. The graph also indicated the complexity of curcumin degradation. The same investigators also investigated the stability of curcumin when exposed to UV/visible radiation (58). The main degradation products were identified. The reaction mechanisms were investigated and the order of the overall degradation reactions and the half-lives of curcumin in different solvents and in the solid state were determined. These workers also examined the photobiological activity of curcumin using bacterial indicator systems (59). On irradiation with visible light, curcumin proved to be phototoxic for Salmonella typhimurium and Escherichia coli, even at very low concentrations. The observed phototoxicity makes curcumin a potential photosensitizing drug, which might find application in the phototherapy of, for example, psoriasis, cancer and bacterial and viral diseases. Recently, the same group (60), complexed curcumin with cyclodextrin to improve the water solubility and the hydrolytic and photochemical stability of the compound. Complex formation resulted in an increase in water solubility at pH 5 by a factor of at least 104. The hydrolytic stability of curcumin under alkaline conditions was strongly improved by complex formation, while the photodecomposition rate was increased compared to a curcumin solution in organic solvents. The cavity size and the charge and bulkiness of the cyclodextrin side-chains influenced the stability constant for complexation and the degradation rate of the curcumin molecule. Wang et al. examined the degradation kinetics of curcumin under various pH conditions and the stability of curcumin in physiological matrices (61). When curcumin was incubated in 0.1 M phosphate buffer and serum-free medium, pH 7.2, at 37ÆC, about 90% decomposed within 30 minutes. A series of pH conditions ranging from 3 to 10 were tested and the result showed that decomposition was pH-dependent and occurred faster at neutral-basic conditions. It is more stable in cell culture medium containing 10% fetal calf serum and in human blood; less than 20% of curcumin decomposed within 1 hour and, after incubation for 8 hours, about 50% of curcumin still remained. Trans-6-(4′-hydroxy-3′-methoxyphenyl)2,4-dioxo-5-hexenal was predicted to be the major degradation product and vanillin, ferulic acid and feruloyl methane were identified as minor degradation products. The amount of vanillin increased with incubation time.

Preclinical Studies
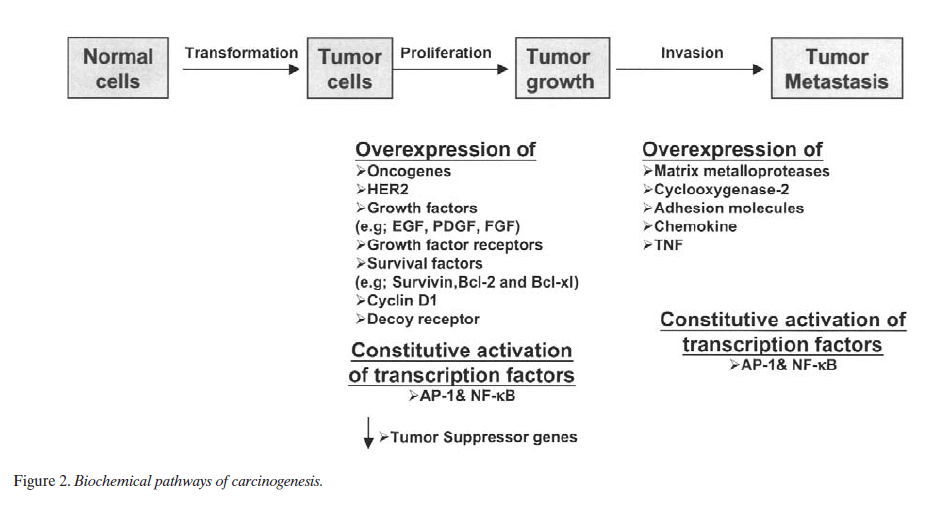
Cancer is a hyperproliferative disorder that metastasize into the vital organs in the body through invasion followed by angiogenesis and distant metastasis. Within the last 25 years, much has been learned about the biochemical pathway that ultimately leads to cancer (Figure 2). During the last decade, there has been extensive investigation of how curcumin affects this overall process of tumorigenesis. Below is the description of the effects of curcumin first in vitro and then in vivo and its relevance to cancer.
In vitro effects
B1A. Antiproliferative effects of curcumin: Curcumin suppresses the proliferation of a wide variety of tumor cells, including breast carcinoma, colon carcinoma, renal cell carcinoma, hepatocellular carcinoma, T cell leukemia, B cell lymphoma, acute myelogenous leukemia, basal cell carcinoma, melanoma and prostate carcinoma (see Table I) (62-93). Additionally curcumin suppresses the proliferation of certain normal cells such as hepatocytes (105), epithelial cells (99), human vascular endothelial cells (HVEC) (96, 97), human vascular smooth muscle cells (HVSMC) (45, 46), osteoclasts (104), peripheral bood mononuclear cells (PBMC) (45), and T lymphocytes (45, 46, 80, 94, 96, 97, 99-105). Curcumin also inhibits the cell proliferation induced by growth factors. For instance IL-2-induced proliferation of PBMC, PDGF-induced proliferation of VSMC and PHA-induced proliferation of PBMC were inhibited by treatment with curcumin (45). The suppression of cell proliferation by curcumin usually occurs through its effects on the cell cycle. Depending on the cell type, the inhibition of cell proliferation at different phases of the cell cycle has been reported (46, 76, 83, 96, 99). Inhibition of cell proliferation could also be mediated through suppression of ornithine decarboxylase (ODC)(65).
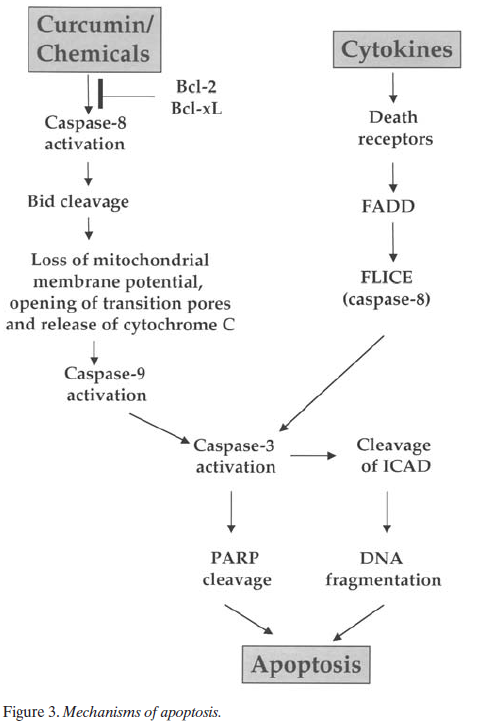
Besides suppression of proliferation, curcumin also induces apoptosis in a wide variety of cells. The mechanism of apoptosis could be either mitochondria-dependent or mitochondria-independent (Figure 3). Normally, cytokines that mediate their signaling through the death receptors induce apoptosis via mitochondria-independent mechanism. Depending on the cell type, curcumin induces apoptosis through both mitochondria-dependent (77, 81-84, 92, 102, 105) as well as mitochondria-independent mechanisms (65, 70, 72). In most cells curcumin sequentially induces activation of caspase-8, cleavage of BID, loss of mitochondrial membrane potential, opening of transition pores, release of cytochrome C, caspase-9 activation, caspase-3 activation and cleavage of PARP and inhibitor of caspase-activated deoxy-ribonuclease (ICAD), thus leading to DNA fragmentation and apoptosis (73, 81, 83, 86, 88, 92, 102). On treatment of cells with curcumin, cell shrinkage, chromatin condensation, translocation of the AIF to the nucleus, nuclear disintegration and large- scale DNA chromatolysis has also been reported (63, 79, 80). Down-regulation of antiapoptotic protein (bcl-2 and bcl-xl) is another potential mechanism by which curcumin could induce apoptosis in cells (46, 64, 74, 83, 84, 86, 88, 92). In breast cancer cells curcumin has been shown to induce bax through p53 which acts on mitochondria, thus leading to apoptosis (91). Curcumin has been shown to induce p53 in colon cancer cells, which could also mediate apoptosis (62). Another potential mechanism through which curcumin could induce apoptosis is through the alteration of the redox status of the cells. Curcumin has been shown to induce ROI and alter cellular GSH levels (64, 77, 86, 87). In contrast to all these studies, there is a recent report that indicates that curcumin could be inhibitory to chemotherapy -induced apoptosis mediated through the suppression of chemotherapy-induced free radical production (93). Inhi-bition of PTK and PKC could also contribute to the ability of curcumin to induce apoptosis. From these studies it is clear that curcumin inhibits cell growth and induces apoptosis of various tumor cells through a mechanism similar to that of most chemotherapeutic agents.

In addition curcumin could affect cellular proliferation through modulation of various cell-signaling pathways (see Table II). These may include transcription factors (e.g, NF-ÎB and AP-1), mitogen-activated protein kinases, growth factor receptor kinases and cyclooxygenases. The mechanism by which curcumin inhibits these pathways is further discussed below. B1B. Curcumin inhibits farnesyl protein transferase (FPTase): Ras proteins must be isoprenylated at a conserved cysteine residue near the carboxyl terminus (Cys- 186 in mammalian Ras p21 proteins) in order to extend their biological activity. Previous studies indicate an intermediate in the mevalonate pathway, most likely farnesyl pyrophosphate, is the donor of this isoprenyl group, and that using inhibitors of the mevalonate pathway could block the transforming properties of ras oncogene. Chen et al. examined the effects of curcumin on farnesyl protein transferase (FTPase) (172). They found that partially purified farnesyl protein transferase (FPTase) capable of catalyzing the farnesylation of unprocessed Ras p21 proteins in vitro was inhibited by curcumin and its derivatives. This is another potential mechanism by which curcumin could suppress cellular growth. B1C. Suppression of NF-κ B activation by curcumin: Members of the NF-κ B transcription factor family play a central role in various responses leading to host defense, activating a rapid progression of gene expression. These transcription factors are dimeric complexes composed of different members of the Rel/NF-κ B family of polypeptides. This family is distinguished by the presence of a Rel homology domain of about 300 amino acids that displays a 35% to 61 % identity between various family members (173). Although NF-κ B is a ubiquitous transcription factor, it plays its critical role in the cells of the immune system, where it controls the expression of various cytokines and the major histocompatibility complex genes. The inappropriate regulation of NF-κ B and its dependent genes have been associated with various pathological conditions including toxic/septic shock, graft vs host reaction, acute inflammatory conditions, acute phase response, viral replication, radiation damage, atherosclerosis, and cancer (173, 174). Unlike other transcription factors, the NF-κ B proteins and other members of the Rel family reside in the cytoplasm in an inactive state; upon activation, they are translocated to the nucleus. The nuclear translocation of Rel proteins is induced by many agents, including inflammatory cytokines (e.g., tumor necrosis factor (TNF), lymphotoxin (LT), and interleukin (IL)-1), mitogens, bacterial products, protein synthesis inhibitors, oxidative stress (H2O2), ultraviolet light and phorbol esters (175, 176). Upon activation of NF-ÎB, a large number of genes are induced, including various inflammatory cytokines (e.g. TNF, IL-1, chemokines), adhesion molecules (e.g. ICAM-1, VCAM I, ELAM-1), COX2, MMP-9 and NOS (175, 176). Because of its intimate involvement in host defense against disease, this transcription factor is an important target for therapeutic intervention. Our laboratory reported that curcumin is a potent inhibitor of NF-κ B activated in response to TNF, PMA and H2O2 (106). The suppression of NF-κ B activation by curcumin was found to be due to the inhibition of Iκ B• degradation, thus abrogating nuclear translocation of p65. These results indicated that curcumin inhibits the NF-κ B activation pathway at a step before IÎB• phosphorylation but after the convergence of various stimuli. Other laboratories have reported that curcumin also suppresses the NF-κ B activated by agents such as IL-1, LPS and chemotherapeutic drugs (108, 110, 115, 122-127, 131, 133, 138, 163, 164, 177-179). More recently, it was demonstrated that curcumin inhibits NF-κ B by inhibiting the kinase that induces the phosphorylation of Iκ B• (Iκ B• kinase; (115, 125, 131,180). The expression of several genes that are regulated by NF-κ B has also been shown to be suppressed by curcumin (80, 107, 111, 112, 119, 121-124, 126, 129, 130, 132, 163-165). These include cell surface adhesion molecules, chemokines, TNF, MMP9, COX2 and NOS. Since these genes are critical regulators of inflammation, the suppression of expression of these genes may explain the anti-inflammatory effects of curcumin. NF-κ B has recently been implicated in the cell survival and proliferation pathways (181, 182), thus the suppression of NF-ÎB may explain the antiproliferative and apoptosis-inducing effects of curcumin. B1D. Suppression of AP-1 by curcumin: The role of various transcription factors in tumorigenesis (tumor initiation, tumor induction and tumor promotion) have been described (183). Activating protein-1 (AP-1) is one such transcription factor transactivated by various tumor-promoting agents, such as phorbol ester, UV radiation, asbestos and crystalline silica (for references see (184, 185)). AP-1 complexes are formed by dimers of Jun proto-oncogene family members (c-Jun, JunB, and JunD) or heterodimers of the Jun family members with the Fos proto-oncogene family members (c-Fos, FosB, Fra-1, and Fra-2). AP-1 binds to a specific target DNA site (also known as the TRE) in the promoters of several cellular genes and mediates immediate early gene expression involved in a diverse set of transcriptional regulation processes (184, 186). Agents that activate NF-κ B also activate the transcription factor AP-1. Both of these factors are regulated by the redox status of the cell. AP-1 activation has been implicated in cell proliferation and in chemical carcinogenesis. Studies in ex vivo and in vivo models showed that the expression of various genes regulated by AP-1 play important roles in the transformation from preneoplastic to neoplastic state (187). AP-1 is also known to be involved in tumor progression and metastasis. Curcumin has been shown to suppress the activation of AP-1 induced by tumor promoters (108, 119, 123, 134, 138, 178). In vitro experiments indicate that inhibition of c-Jun/AP-1 binding to its cognate motif by curcumin may be responsible for the inhibition of c-Jun/AP-1-mediated gene expression. The expression of several AP-1-regulated genes has been shown to be regulated by curcumin. These include c-fos, c-jun, c-myc, endothelial tissue factor, chemokines and MMP (107, 119, 123, 135-137). The mechanism by which curcumin inhibits AP-1 is not fully understood, but there are three potential mechanisms: first through alteration of the redox status of the cells; second, through inhibition of JNK (188), a kinase needed for AP-1 activation; and third, through inhibition of the fos-jun-DNA complex (189). Thus down-regulation of AP-1 by curcumin may explain its ability to suppress chemical carcinogenesis. B1E. Suppression of Egr-1 by curcumin: The transcription factor early growth response-1 gene product (Egr-1) is a member of the family of immediate early response genes and regulates a number of patho-physiologically relevant genes in vasculature that are involved in growth, differentiation, immune response, wound healing and blood clotting. Pendurthi et al. investigated the effect of curcumin on Egr-1 expression in endothelial cells and fibroblasts (140). Gel mobility shift assays showed that pretreatment of endothelial cells and fibroblasts with curcumin suppressed TPA and serum-induced Egr-1 binding to the consensus Egr-1 binding site and also to the Egr-1 binding site present in the promoter of the tissue factor gene. Western blot analysis revealed that curcumin inhibited TPA-induced de novo synthesis of Egr-1 protein in endothelial cells. Suppression of Egr-1 protein expression in curcumin-treated cells stemmed from the suppression of Egr-1 mRNA. Northern blot analysis showed that curcumin inhibited serum and TPA-induced expression of tissue factor and urokinase-type plasminogen activator receptor mRNA in fibroblasts. These results showed that curcumin suppresses the induction of Egr-1 and thereby modulates the expression of Egr-1-regulated genes in endothelial cells and fibroblasts. The down-regulation of tissue factor by curcumin has also been demonstrated by another group (123). B1F. Suppression of mitogen-activated protein kinases by curcumin: Most inflammatory stimuli are known to activate three independent mitogen-activated protein kinase (MAPK) pathways, leading to activation of p44/42 MAPK (also called ERK1/ERK2), JNK and p38 MAPK pathway. Chen et al. found that curcumin inhibits JNK activation induced by various agonists including PMA plus ionomycin, anisomycin, UV-C, gamma radiation, TNF and sodium orthovanadate (188). Although both JNK and ERK activation by PMA plus ionomycin were suppressed by curcumin, the JNK pathway was more sensitive. The IC50 (50% inhibition concentration) of curcumin was between 5-10 ÌM for JNK activation and was 20 ÌM for ERK activation. In transfection assays, curcumin moderately suppressed MEKK1-induced JNK activation; however, it effectively blocked JNK activation caused by co-transfection of TAK1, GCK, or HPK1. Curcumin did not directly inhibit JNK, SEK1, MEKK1 or HPK1 activity. Although curcumin suppressed TAK1 and GCK activities at high concentrations, this inhibition cannot fully account for the JNK inhibition by curcumin in vivo. Thus these results suggested that curcumin affected the JNK pathway by interfering with the signaling molecule(s) at the same level or proximally upstream of the MAPKKK level. The inhibition of the MEKK1-JNK pathway reveals a possible mechanism of suppression of AP-1 and NF-κ B signaling by curcumin and may explain the potent anti-inflammatory and anti-carcinogenic effects of this chemical. B1G. Suppression protein kinases by curcumin: Curcumin could also mediate its effects through inhibition of various other serine/threonine protein kinases. Our group showed that treatment of highly purified protein kinase A (PKA), protein kinase C (PKC), protamine kinase (cPK), phospho-rylase kinase (PhK), autophosphorylation-activated protein kinase (AK), and pp60c-src tyrosine kinase with curcumin inhibited all kinases. PhK was completely inhibited at low concentration of curcumin (147). At around 0.1 mM curcumin, PhK, pp60c-src, PKC, PKA, AK and cPK were inhibited by 98%, 40%,15%, 10%, 1% and 0.5%, respectively. Lineweaver-Burk plot analysis indicated that curcumin is a noncompetitive inhibitor of PhK with a Ki of 0.075 mM. Other investigators have shown suppression of PMA-induced activation of cellular PKC by curcumin (146). Treatment of cells with 15 or 20 ÌM curcumin inhibited TPA-induced PKC activity in the particulate fraction by 26% or 60%, respectively, and did not affect the level of PKC. Curcumin also inhibited PKC activity in both cytosolic and particulate fractions in vitro by competing with phospha-tidylserine. However, the inhibitory effect of curcumin was reduced after preincubation with the thiol compounds. These findings suggested that the suppression of PKC activity may contribute to the molecular mechanism of inhibition of TPA-induced tumor promotion by curcumin. Besides in vitro suppression, curcumin could also inhibit PKC in the cells (148). Hasmeda et al. showed that curcumin inhibits Ca2+-and phospholipid-dependent PKC and of the catalytic subunit of cyclic AMP-dependent protein kinase (cAK; IC50 values 15 and 4.8 ÌM, respectively) (148). Curcumin inhibits plant Ca2+-dependent protein kinase (CDPK) (IC50 41 ÌM), but does not inhibit myosin light chain kinase or a high-affinity 3′,5′-cyclic AMP-binding phosphatase. Curcumin inhibits cAK, PKC and CDPK in a fashion that is competitive with respect to both ATP and the synthetic peptide substrate employed. The IC50 values for inhibition of cAK by curcumin are very similar when measured with kemptide (LRRASLG) (in the presence or absence of ovalbumin) or with casein or histone III-S as substrates. However, the presence of bovine serum albumin (0.8 mg ml-1) largely overcomes inhibition of cAK by curcumin. µ1∏ Curcumin inhibits growth factor receptor protein tyrosine kinases: Besides serine protein kinases, curcumin has also been shown to inhibit protein tyrosine kinase activity (PTK) of the EGF receptor (142, 143, 145). Korutla et al. showed that the short term treatment of cells with curcumin inhibited EGF receptor intrinsic kinase activity by up to 90%, and also inhibited EGF-induced tyrosine phosphorylation of EGF receptors (142). Another study found that the treatment of cells with a saturating concentration of EGF for 5-15 minutes induced increased EGF-R tyrosine phosphorylation, and this induction was inhibited by up to 90% by curcumin, which also inhibited the growth of EGF-stimulated cells (143). Curcumin treatment had no effect on the amount of surface expression of labeled EGF-R, and inhibition of EGF-mediated tyrosine phosphorylation of EGF-R by curcumin was mediated by a reversible mechanism. In addition, curcumin also inhibited EGF-induced, but not bradykinin-induced, calcium release. These findings demonstrate that curcumin is a potent inhibitor of a growth stimulatory pathway, the ligand-induced activation of EGF-R, and may be useful in developing anti – proliferative strategies to control tumor cell growth. The erbB2/neu gene-encoded p185neu tyrosine kinase is a potent oncoprotein. Overexpression of p185neu in breast cancer is known as a prognostic factor. Hong et al. investigated the effect of curcumin on p185neu tyrosine kinase and on the growth of breast cancer cell lines (144). Curcumin inhibited p185neu autophosphorylation and transphosphorylation in vitro and depleted p185neu protein in vivo. It dissociated the binding of p185neu with GRP94 (glucose-regulated protein), a molecular chaperone, and enhanced the depletion of p185neu. The amount of p185neu protein on the cell membrane was drastically decreased after curcumin treatment. These data demonstrated a new mechanism of the anti-tyrosine kinase activity of curcumin. The growth of several breast cancer cell lines was inhibited; the IC50 ranged from 7 to 18 ÌM, which, however, did not correlate with the expression level of p185neu. Colony formation in the soft agar assay, a hallmark of the transformation phenotype, was preferentially suppressed in p185neu-overexpressing cell lines by 5 ÌM curcumin. Because curcumin effectively inhibited p185neu tyrosine kinase activity by depleting p185neu and potently suppressed the growth of multiple breast cancer cell lines, its therapeutic potential in advanced breast cancer is worthy of further investigation. Dorai and coworkers examined the effect of curcumin on EGF receptor signaling in the androgen- sensitive LNCaP and androgen-insensitive PC-3 cell lines (145). They found that curcumin was a potent inhibitor of EGF-R signaling, and it accomplished this effect by three different means: (1) down-regulating the EGF-R protein; (2) inhibiting the intrinsic EGF-R tyrosine kinase activity; and (3) inhibiting the ligand-induced activation of the EGF-R. They concluded that curcumin can induce apoptosis by interfering with the signal transduction pathways of the prostate cancer cell. B1 π. Suppression of COX2 and lipoxygenase (LOX) expression by curcumin: COX2 and LOX are important enzymes that mediate inflammation through production of prostaglandins and leukotrienes, respectively. Curcumin has been shown to suppress the expression of both COX2 and LOX protein as well as their enzymatic activities. Huang et al., (134,149) showed that topical application of curcumin markedly inhibited TPA- and arachidonic acid-induced epidermal inflammation (ear edema) in mice. The in vitro addition of curcumin to cytosol from homogenates of mouse epidermis inhibited the metabolic conversion of arachidonic acid to 5-hydroxyeicosatetraenoic acid (5-HETE), of arachidonic acid to 8-HETE, and of arachidonic acid to prostaglandin (PG) E2, PGF2•, and PGD2. The inhibitory effects of curcumin on TPA-induced tumor promotion in mouse epidermis paralleled their inhibitory effects on TPA-induced epidermal inflammation and epidermal LOX and COX activities. Ammon et al. showed that curcumin acts as an anti-inflammatory in in vivo animal model sthrough inhibition of 5-LOX and 12-LOX activity in rat peritoneal neutrophils and COX activity in human platelets (190). Zhang et al. investigated whether curcumin inhibits chenodeoxycholate – or PMA-mediated induction of COX-2 in several gastrointestinal cell lines (124). Treatment with curcumin suppressed chenodeoxycholate – and PMA-mediated indu-ction of COX2 protein and synthesis of PGE2. It also suppressed the induction of COX-2 mRNA by cheno-deoxycholate and PMA. Nuclear run-offs revealed increased rates of COX2 transcription after treatment with cheno-deoxycholate or PMA, and these effects were inhibited by curcumin. Treatment with chenodeoxycholate or PMA increased binding of AP-1 to DNA. This effect was also blocked by curcumin. In addition to the above effects on gene expression, Zhang et al. found that curcumin directly inhibited the expression of COX2 (124). Ramsewak and his group examined curcumin π, curcumin ππ (demethoxycurcumin) and curcumin III (bisdemetho-xycurcumin) for their cytotoxicity and antioxidant and anti-inflammatory activities (95). These compounds were active against leukemia, colon, CNS, melanoma, renal and breast cancer cell lines. Curcumins π, ππ and III at 100 Ìg/ml inhibited liposome peroxidation by 58%, 40% and 22%, respectively. Curcumins π, ππ and III were active against COX1 enzyme at 125 Ìg/ml and showed 32%, 38% and 39% inhibition of the enzyme, respectively. Curcumins π, ππ, and πππ at 125 Ìg/ml inhibited the COX-ππ enzyme by 90%, 82% and 59% inhibition of the enzyme, respectively. Plummer et al. also examined the effect of curcumin on COX2 levels in colorectal cancer (125). They found that curcumin inhibited COX2 induction by the colon tumor promoters TNF or fecapentaene-12 through inhibition of NF-ÎB. Similar results were reported by Goel et al. in colon cancer cells (126). Surh et al. found that NF-κ B is involved in the regulation of COX2 expression by curcumin (127). Skrzypczak-Jankun et al. discovered a novel mechanism for inhibition of LOX by curcumin (150). They investigated the fate of curcumin when used as a soybean lipoxygenase L3 substrate. By use of X-ray diffraction and mass spectrometry, they found that curcumin can act as a LOX substrate: specifically, an unoccupied electron mass that appeared to be an unusual degradation product of curcumin (4-hydro-xyperoxy-2-methoxyphenol) was located near the soybean L3 catalytic site. Understanding how curcumininhibits LOX may help in the development of novel anti-cancer drugs. B1J. Suppression of cyclin D1 by curcumin: Cyclins are essential for cell division, activating their partners, the cyclin-dependent kinases, and directing them to specific substrates. One of the subgroup of cyclins, called cyclin D, consists of three known subtypes, D1, D2 and D3, all of which collectively control cell cycle progression by activating their cyclin-dependent kinase partners, CDK4 and CDK6. These kinases then phosphorylate the retinoblastoma protein and thus advance the cell through the G1-phase of the cell cycle (191, 192), leading to stimulation of DNA synthesis. (192) Cyclin D1 is a proto-oncogene that is overexpressed as a result of gene amplification or translocation in many cancers, including breast, esophagus, lung, liver, head and neck, colon and prostate (193-203). For instance, the cyclinD1 gene is amplified in 20-50% of squamous cell carcinoma (SCC), and its protein is overexpressed in up to 80% of SCCs (198) . Overexpression of cyclin D1 is also associated with metastatic prostate cancer to bone (202, 203). The cyclin D1 gene is also amplified in up to 20% of human breast cancers (197), while cyclin D1 protein is overexpressed in some 50% of human breast cancers (194, 195) and is required for the proliferation of breast cancer cells in culture (204, 205). Transgenic mice engineered to overexpress cyclin D1 in mammary glands develop breast cancer, thus suggesting a causative role for this protein (205). Cyclin D1 also has been associated with aggressive forms of hepatocellular carcinoma (196, 206). The genetic deletion of cyclin D1 in mice impaired breast development during pregnancy (207-210), and these mice were found to be resistant to breast cancers induced by Ras, rac and neu oncogenes (211-213). In comparison, genetic deletion of cyclin D2 and D3 had no effect. Thus tumor cell proliferation by Ras and Neu in breast cancer cells is mediated through cyclin D1 only. Furthermore, Ras and Neu have been shown to regulate the promoter region of the cyclin D1 gene (211, 213). Thus drugs that can down-regulate cyclin D1 have potential as treatments for breast cancer and other tumors. Antisense to cyclin D1 has been used to down-regulate cyclin D1 and shown to induce apoptosis and tumor shrinkage in SCC (214). Our laboratory found that inhibition of the proliferation of prostate cancer, breast cancer and multiple myeloma cell lines by curcumin correlated with the down-regulation of the expression of cyclin D1 protein (180, 215). The suppression of cyclin D1 by curcumin led to inhibition of CDK4-mediated phosphorylation of retinoblastoma protein. Curcumin-inducd down-regulation of cyclin D1 was inhibited by lactacystin, an inhibitor of 26S proteosome, suggesting that curcumin represses cyclin D1 expresion by promoting proteolysis. Curcumin also down-regulated mRNA expression and inhibited the activity of cyclin D1 promoter-dependent reporter gene expression. Thus curcumin down-regulates cyclin D1 expression through activation of both transcri-ptional and post-transcriptional mechanisms, and this may contribute to the antiproliferative effects of curcumin. B1K. Suppression of adhesion molecules by curcumin: Recruitment of leukocytes by endothelial cells (EC) and their subsequent migration is known to play a critical role in inflammation. TNF is a multifunctional cytokine shown to be involved in inflammation through the expression of various inflammatory molecules (216). It induces the expression of adhesion molecules on EC involved in transendothelial cell migration of leukocytes (217-219). In particular TNF has been shown to induce de novo synthesis of intracellular adhesion molecule-1 (ICAM-1, also called CD54), vascular cell adhesion molecule-1 (VCAM-1), and endothelial leukocyte adhesion molecule-1 (ELAM-1, also called E-selectin). ICAM-1 and VCAM-1 are 95-kDa and 110-kDa proteins, respectively, and both belong to the immunoglobulin superfamily. Besides EC, ICAM-1 is also expressed by monocytes, B and T cells, keratinocytes, chondrocytes and epithelial cells. In addition to EC, monocytes, and dendritic cells, VCAM-1 is also expressed in myoblast and bone marrow fibroblasts. The 115-kDa protein ELAM-1, which belongs to the selectin family, is expressed exclusively on EC. We investigated the effect of curcumin on adhesion of monocytes to human umbilical vein endothelial cells (EC) (121). Treatment of endothelial cells with tumor necrosis factor (TNF) augmented the adhesion of monocytes to endothelial cells, and this adhesion was due to increased expression of ICAM-1, VCAM-1 and ELAM-1. Curcumin completely blocked the adhesion of monocytes with endothelial cells as well as the cell surface expression of ICAM-1, VCAM-1 and ELAM-1 in EC. That curcumin inhibits TNF-induced expression of adhesion molecules on human umbilical vein endothelial cells has also been reported by others (122). Although curcumin inhibited adhesion even when administered 1 hour after TNF treatment, maximum inhibition occurred when added either 1 hour before or at the same time as TNF. The inhibition of TNF-induced adhesion molecules was found to be due to abrogation of NF-κ B, activation. Our results demonstrated that curcumin’s anti-inflammatory properties may be attributable in part to inhibition of leukocyte recruitment. Since cell adhesion molecules play a determining role in the tumor metastasis, their down-regulation by curcumin may contribute to its anti-cancer properties. B1L. Suppression of Ca2+ ATPase by curcumin: Logan-Smith et al. found that curcumin is a potent inhibitor of the Ca2+-ATPase of sarcoplasmic reticulum (SR) but increases the rate of accumulation of Ca2+ (158). Curcumin is an inhibitor of the ATPase activity of the Ca2+-ATPase of skeletal muscle SR. Inhibition by curcumin is structurally specific, requiring the presence of a pair of -OH groups at the 4-position of the rings. Inhibition is not competitive with ATP. Unexpectedly, addition of curcumin to SR vesicles leads to an increase in the rate of accumulation of Ca2+, unlike other inhibitors of Ca 2+-ATPase which reduce the rate of accumulation. An increase in the rate of accumulation of Ca2+ is seen in the presence of phosphate ion, which lowers the concentration of free Ca2+ within the lumen of the SR, showing that the effect is not a passive leak across the SR membrane. The effect is to reduce the rate of slippage on the ATPase, a process in which a Ca2+-bound, phosphorylated intermediate releases its bound Ca2+ on the cytoplasmic rather than on the lumenal side of the membrane. The structural specificity of the effects of curcumin on ATPase activity and on Ca2+ accumulation is the same, and the apparent dissociation constants for the two effects are similar, suggesting that the two effects of curcumin could follow from binding to a single site on the ATPase. B1M. Suppression of production of inflammatory cytokine by curcumin: Curcumin has been found to suppress the expression of several inflammatory cytokines including TNF, IL-1, IL-12 and chemokines (107, 111-113, 163). Chen et al. showed that curcumin, at 5 ÌM, inhibited lipopolysaccharide (LPS)-induced production of TNF and IL-1 by a human monocytic macrophage cell line, Mono Mac 6 (111). In addition, curcumin inhibited LPS-induced activation of NF-κ B and reduced the biological activity of TNF in L929 fibroblast lytic assay. Hanazawa et al. showed that TNF stimulates the expression of the monocyte chemoattractant JE (MCP-1 /JE) gene in osteoblastic MC3T3-E1 cells (135). TNF stimulated this MCP-1/JE gene expression trans-criptionally through expression of the early protooncogenes c-fos and c-jun. Curcumin, markedly inhibited JE gene expression and monocyte chemotactic activity induced by the cytokine. Xu et al. investigated the effect of curcumin on the expression of MCP-1/JE and interferon inducible protein-10kDA (IP-10) in mouse bone marrow stromal cell line +/+-1.LDA11 (107). Both chemokines were readily expressed in stromal cells after stimulation IL-1•, IFN-Á, TNF-• and LPS. Curcumin attenuated the levels of MCP-1/JE and IP-10 mRNA expression by all of these stimulatory agents. Curcumin inhibited both chemokine mRNAs in a dose- and time-dependent manner. The suppressive effect of curcumin on both mRNAs was reversible with complete recovery from suppression within 24 hours after removal of curcumin. The suppression of mRNA by curcumin was dependent on de novo synthesis of an intermediary protein(s), since suppression was abrogated by concomitant treatment with cycloheximide (CHX). Destabilization of mRNA transcripts was not the mechanism by which curcumin lowered the levels of mRNA; however, transcripts formed in the presence of curcumin were more stable, as indicated by their slower degradation kinetics. Run-on transcriptional assays demonstrated that curcumin inhibits the transcriptional activity of both genes. The attenuation of chemokine gene expression was associated with decreased production of chemotactic activity. These findings indicate that while curcumin may post-transcriptionally stabilize mRNA transcripts formed in its presence, the overall reduction in mRNA levels by curcumin is mediated by inhibition of the transcription of chemokine genes (135). Abe et al. investigated the effect of curcumin on inflammatory cytokine production by PBMC and alveolar macrophages (113). They examined the levels of IL-8, monocyte inflammatory protein-1 (MIP-1•), MCP-1, IL-10 and TNF in the culture supernatants from PMA- or LPS-stimulated monocytes and alveolar macrophages in the presence or absence of curcumin. Curcumin inhibited the production of IL-8, MIP-1 MCP-1, IL-10 and TNF by PMA-or LPS-stimulated monocytes and alveolar macrophages. Literat et al. also examined the regulation of pro-inflammatory cytokine expression by curcumin in hyaline membrane disease (HMD) (114). Persistent expression of pro-inflammatory cytokines is believed to play a major role in the pathogenesis of chronic lung disease (CLD) in premature infants. Inhibition of pro-inflammatory cytokine production in the lungs of preterm newborns may result in the attenuation of CLD. Literat’s group derived lung inflamma-tory cells from preterm newborns at risk for the development of CLD via modified broncho-alveolar lavage and stimulated them ex vivo with LPS (10 ng/ml). Curcumin was added to these cultures at 0, 0.5 and 20 ÌM concentrations. Pro-inflammatory cytokine, TNF, IL-10 and IL-8 protein from the culture supernatants were measured 12 hours after culture. For control, PBMC were cultured under the same conditions. Both neonatal lung inflammatory cells and adult PBMC produced high levels of pro-inflammatory cytokines in response to LPS. Curcumin significantly inhibited IL-10 and IL-8 but minimally inhibited TNF expression by preterm lung inflammatory cells at 20 ÌM concentration. Adult PBMC expression of IL-8 was significantly inhibited by curcumin at a 20 ÌM concentration. Therefore, curcumin inhibits pro-inflammatory cytokine production (TNF, IL-1ß and IL-8) by lung inflammatory cells ex vivo. Kang et al. examined the effect of curcumin on IL-12 production from mouse macrophages stimulated with LPS (164). Curcumin potently inhibited the production of IL-12, The effect of curcumin on IL-12 p40 promoter activation was analyzed by transfecting RAW264.7 monocytic cells with p40 promoter/reporter constructs. The repressive effect mapped to a region in the p40 promoter containing a binding site for NF-κ B (p40-κ B). Furthermore, activation of macrophages by LPS resulted in markedly enhanced binding activity to the κ B site, which significantly decreased upon addition of curcumin. The same group (163) also examined the effect of curcumin on IL-12 production by mouse splenic macrophages and the subsequent ability of these cells to regulate cytokine production by CD4+ T cells. Pretreatment with curcumin significantly inhibited IL-12 production by macrophages stimulated with either LPS or heat-killed Listeria monocytogenes (HKL). Curcumin pretreated macrophages reduced their ability to induce IFN-Á and increased the ability to induce IL-4 in Ag-primed CD4+ T cells. Addition of recombinant IL-12 to cultures of curcumin-pretreated macrophages and CD4+ T cells restored IFN-Á production in CD4+ T cells. The in vivo administration of curcumin resulted in the inhibition of IL-12 production by macrophages stimulated in vitro with either LPS or HKL, leading to the inhibition of Thl cytokine profile (decreased IFN-Á and increased IL-4 production) in CD4+ T cells. These findings suggest that curcumin may inhibit the Th 1 cytokine profile in CD4+ T cells by suppressing IL-12 production in macro-phages and point to a possible therapeutic use of curcumin in Th 1 -mediated immune diseases. Natarajan et al. found that curcumin inhibits experimental allergic encephalomyelitis (EAE) in association with a decrease in IL-12 production from macrophage/microglial cells and differentiation of neural Ag-specific Th1 cells (24). In vitro treatment of activated T cells with curcumin inhibited IL-12-induced tyrosine phosphorylation of Janus kinase 2, tyrosine kinase 2, and STAT3 and STAT4 transcription factors. The inhibition of Janus kinase-STAT pathway by curcumin decreased IL-12-induced T cell proliferation and Th1 differentiation. These findings highlight the fact that curcumin inhibits EAE by blocking IL-12 signaling in T cells and suggest its use in the treatment of multiple sclerosis and other Th1 cell-mediated inflammatory diseases. Thus, overall these studies suggests that cucumin has potent immuno-suppressive effects. B1N. Suppression of angiogenesis by curcumin: Angiogenesis is a crucial step in the growth and metastasis of many cancers. The effect of curcumin on endothelial cell migration, attachment, and tube formation on Matrigel has been investigated (220). Curcumin had no effect on endothelial cell migration or attachment to either plastic or Matrigel. Curcumin inhibited tube formation and also caused the preformed tubes to break down. Curcumin inhibited angioge-nesis in a subcutaneus Matrigel plug model in mice. The study also showed that curcumin inhibits the gelatinolytic activities of secreted 53- and 72-kDa MMP and suppresses the expression and transcription of the 72kDa, MMP, indicating its inhibitory effect at both the transcriptional and post-transcriptional level, thus suggesting that curcumin acts as an angiogenesis inhibitor by modulating MMP. Cell motility is essential for a wide range of cellular activities including anigogenesis. In the highly invasive SK-Hep-1 cell line of human hepatocellular carcinoma (HCC), Lin et al. found that curcumin inhibited cellular migration and invasion of SK-Hep-1 (117). Further, and parallel with its anti-invasion activity, curcumin inhibited MMP-9 secretion in SK-Hep-1. Shin et al. studied the role of JNK in endothelial cell motility using stable transfectant (DAR-ECV) of ECV304 endothelial cells expressing previously established oncogenic H-Ras (leu 61) and used curcumin to inhibit it (221). DAR-ECV cells showed an enhanced angiogenic potential and motility (approximately 2-fold) compared to ECV304 cells. Western blot analysis revealed constitutive activation of JNK in DAR-ECV cells. Pretreatment of curcumin decreased the basal motility of DAR-ECV cells in a dose-dependent manner and suppressed the motility stimulated by known JNK agonists such as TNF• and anisomycin. These results suggest that the JNK pathway regulates the motility of endothelial cells during angiogenesis and curcumin can inhibit it. Arbiser et al. investigated Rcurcumin for its ability to inhibit the proliferation of primary endothelial cells in the presence and absence of basic fibroblast growth factor (bFGF), as well as its ability to inhibit proliferation of an immortalized endothelial cell line (222). They found that curcumin effectively inhibited endothelial cell proliferation, inhibited bFGF-mediated corneal neovascularization in the mouse and had no effect on phorbol ester-stimulated VEGF production. These results indicate that curcumin has direct antiangiogenic activity in vitro and in vivo. Using cultured corneal cells, Mohan et al. showed that FGF -2 stimulates DNA binding activity of AP-1 but not NF -κ B and that AP-1 stimulation is inhibited by curcu-minoids (118). The FGF-2-induced gelatinase B trans-criptional promoter activity was found to be dependent on AP-1 but not NF-κ B response elements, and promoter activity was also inhibited by curcuminoids. In rabbit corneas, the angiogenic response induced by implantation of an FGF-2 pellet was inhibited by the co-implantation of a curcuminoid pellet and this correlated with inhibition of endogenous gelatinase B expression induced by FGF-2. Angiostatic efficacy in the cornea is also observed when curcuminoids were provided to mice in the diet. Thus these results also provide evidence that curcuminoids target the FGF-2 angiogenic signaling pathway and inhibit expression of gelatinase B in the angiogenic process. TGF-ß1 stimulates migration/invasion of mouse trans-formed keratinocytes and increases urokinase (uPA) expression/secretion. Santibanez et al. found that curcumin abrogated the enhancement of uPA levels induced by TGF-ß1 in transformed keratinocytes; inhibited the TGF-ß1-induced synthesis of fibronectin, an early response gene to the growth factor; and reduced TGF-ß1-stimulated cell migration and invasiveness (165). These results suggest that a tyrosine kinase- signaling pathway should be involved in TGF-ß1-mediated increase in malignancy of transformed keratinocytes and that curcumin could play an important role in inhibiting this process. B1O. Curcumin inhibits xanthine oxidase: Lin et al. showed that the treatment of cells with the PMA elevates xanthine oxidase (XO) activity, an enzyme capable of generating reactive oxygen species such as superoxide and hydrogen peroxide (154). Simultaneous administration of 2 and 10 ÌM curcumin with 100 ng/ml PMA inhibited PMA-induced increases in XO activity. The PMA-induced conversion of xanthine dehydrogenase (XD) to XO was reduced by curcumin to the basal level noted in untreated cells. The activity of XO is remarkably inhibited by curcumin in vitro, but not by its structurally related compounds caffeic acid, chlorogenic acid and ferulic acid. Based on these findings, induction of XO activity is thought to be one of the major causes of PMA-mediated tumor promotion, and the major mechanism by which curcumin inhibits PMA-induced increases in XD/XO enzyme activities is through direct inactivation at the protein level. B1P. Curcumin activates aryl hydrocarbon receptor and induces cytochrome P450: Various carcinogens activate the pathway mediated by the aryl hydrocarbon receptor (AhR). Ciolino et al. examined the effect of curcumin on the AhR and cytochrome P450 1A1 in MCF-7 cells. curcumin caused a rapid accumulation of cytochrome P450 1A1 (CYP1A1) mRNA, and CYP1A1 monooxygenase activity increased as measured by ethoxyresorufin – O – deethylation (157). Curcumin activated the DNA- binding capacity of the AhR for the xenobiotic responsive element of CYP1A1 as measured by electrophoratic mobility shift assay. Curcumin was able to compete with the prototypical AhR ligand 2,3,7,8-tetrachlorodibenzo-p-dioxin for binding to the AhR in isolated MCF-7 cytosol, indicating that it interacts directly with the receptor. Although curcumin could activate the AhR on its own, it partially inhibited the activation of AhR, as measured by electrophoratic mobility shift assay, and partially decreased the accumulation of CYP1A1 mRNA caused by the mammary carcinogen dimethylbenzanthracene (DMBA). Curcumin compe-titively inhibited CYP1A1 activity in DMBA-treated cells and in microsomes isolated from DMBA-treated cells. Curcumin also inhibited the metabolic activation of DMBA, as measured by the formation of DMBA-DNA adducts, and decreased DMBA-induced cytotoxicity. These results suggest that the chemopreventive effect of curcumin may be due, in part, to its ability to compete with aryl hydrocarbons for both the AhR and CYP1A1 sites. Curcumin may thus be a natural ligand and substrate of the AhR pathway. Oetari et al. investigated the interactions between curcumin and cytochrome P450s (P450s) in rat liver (152). Curcumin is relatively unstable in phosphate buffer at pH 7.4. The stability of curcumin was strongly improved by lowering the pH or by adding glutathione (GSH), N-acetyl L-cysteine (NAC), ascorbic acid, rat liver microsomes, or rat liver cytosol. Curcumin was found to be a potent inhibitor of rat liver P450 lAl/lA2 measured as ethoxyresorufin deethylation (EROD) activity in ß-naphthoflavone (ßNF)-induced microsomes, a less potent inhibitor of P450 2B1/2µ2, measured as pentoxyresorufin depentylation (PROD) activity in pheno-barbital PB-induced microsomes, and a weak inhibitor of P450 2EI, measured as p-nitrophenol (PNP) hydroxylation activity in pyrazole-induced microsomes. Ki values were 0.14 and 76.02 ÌM for the EROD and PROD activities, respe-ctively, and 30 ÌM curcumin inhibited PNP-hydroxylation activity by only 9%. In EROD and PROD experiments, curcumin showed a competitive type of inhibition. B1Q. Curcumin binds to P-glycoprotein and induces chemo-sensitivity: Curcumin has been endowed with beneficial biological activities, including antioxidant, anticarcinogenic and hepatoprotective effects. Romiti et al. examined the effects of curcumin on P-glycoprotein in primary cultures of rat hepatocytes for its potential ability to interact in vitro with hepatic P-glycoprotein (Pgp) (169). In this system, sponta-neous overexpression of multidrug resistance (mdr) genes occurs. In both freshly-plated hepatocytes, containing low levels of Pgp, and 72 hour-cultured hepatocytes, containing high levels of Pgp, the rhodamine-123 (R-123) efflux, which represents a specific functional test for Pgp-mediated transport, was inhibited by curcumin. A 25 ÌM dose of curcumin significantly lowered the increase of mAb C219-immunoreactive protein that spontaneously occurs in cells during culture. Curcumin, at doses ranging from 50 to 150 ÌM, was cytotoxic for freshly plated hepatocytes, as shown by the strong decrease in the cell’s ability to exclude trypan blue 24 hours later, but it was significantly less cytotoxic when added to cells cultured for 24 or 48 hours. The resistance to curcumin, progressively acquired by cells during culture was significantly reduced by high concentrations of dexa-methasone (DEX) or DMSO, culture conditions known to inhibit the spontaneous overexpression of Pgp. In addition, in a concentration-dependent manner, verapamil reverted curcumin resistance in Pgp overexpressing hepatocytes. In photoaffinity labeling studies, curcumin competed with azidopine for binding to Pgp, suggesting a direct interaction with glycoprotein. Recently, Anuchapreeda et al. also studied the effect of curcumin on the expression and function of Pgp in the multidrug-resistant human cervical carcinoma cell line ∫µ-V1 (223). Curcumin lowered Pgp expression and MDRI mRNA levels in ∫µ-V1 cells in a concentration-dependent manner. The effect of curcumin on Pgp function was demonstrated by rhodamine 123 (Rh123) accumulation and efflux in Pgp-expressing ∫µ-V1 cells. Curcumin increased Rh123 accumulation in a concentration-dependent manner (1 -55 microM) and inhibited the efflux of Rh123 from these cells, but did not affect the efflux of Rh123 from the wild-type drug-sensitive ∫µ-3-1 cells. Treatment of drug-resistant ∫µ-V1 cells with curcumin increased their sensitivity to vinblastine, which was consistent with an increased intracellular accumulation of Rh123. In addition, curcumin inhibited verapamil-stimulated ATPase activity and the photoaffinity labeling of Pgp with the prazosin analog [125π] iodoarylazidoprazosin in a concentration-dependent manner, demonstrating that curcumin interacts directly with the transporter. These results suggest that in vitro curcumin is able to modulate both expression and function of Pgp. Thus, curcumin could also reveal itself to be a compound endowed with chemosensitizing properties in cells with the mdr phenotype. B1R. Curcumin interacts and inhibits glutathione s-transferase: Anticarcinogenic, antimutagenic, antioxidant and cytopro-tective effects of curcumin can be explained by its inhibitory effect on glutathione s-transferase (GST). Susan et al. studied the induction of GST activity by curcumin in mice (15 1). At a dose of 250 mg/kg orally for 15 days, the enzyme activity in liver was increased by 1.8-fold. Its effect on the stomach, small intestine, lungs and kidney was not significant. curcumin also depleted sulfhydryl levels in tissues, especially in the stomach, where 45% depletion was observed. Oetari et al. investigated the interactions between curcumin and GST in rat liver (152). curcumin is relatively unstable in phosphate buffer at pH 7.4, but the stability was strongly improved by lowering the pH or by adding glutathione (GSH), N-acetyl L-cysteine (NAC), ascorbic acid, rat liver microsomes, or rat liver cytosol. Curcumin was found to be a potent inhibitor of GST activity in cytosol from liver of rats treated with phenobarbital (PB), Pnaphthoflavone (PNF) and pyrazole (Pyr), when measured using 1-chloro-2,4-dinitrobenzene (CDNB) as substrate. In liver cytosol from rats treated with PB, curcumin inhibited GST activity with Ki of 5.75 ÌM and 12.5 ÌM. In liver cytosol from rats treated with pyrazole (Pyr) or beta-naphthoflavone (beta NF), curcumin demonstrated a competitive type of inhibition with Ki values of 1.79 ÌM and 2.29 ̪, respectively. πersel et al. reported that curcumin inhibits GST activity towards 1-chloro-2,4dinitrobenzene in human melanoma cells (224). The major GST subunit expressed in these cells is the pi-class GST subunit P1. A 1-hour exposure of GST-treated cells to 25 ÌM curcumin caused 96% inhibition. Up to about 50% GSH-depletion was found after treatment with curcumin. Curcumin inactivated GSTP1-1 by covalent modi-fication. This was clear from the fact that, depending on the dose, between 30% and 80% inhibition was still observed after lysis of the cells, under which conditions inhibition is no longer reversible. Awasthi et al. also examined the interaction of glutathione (GSH) with curcumin (153). Curcumin contains two ele-ctrophilic •, ß-unsaturated carbonyl groups, which can react with nucleophilic compounds such as GSH. Awasthi’s group took advantage of this characteristic in investigating the reactions of curcumin with GSH and the effect of recombinant human GSTP1-1 on reaction kinetics. Gluta-thionylated products of curcumin identified by FAB-MS and MALDI-MS included mono- and di-glutathionyl adducts of curcumin as well as cyclic rearrangement products of GSH adducts of feruloylmethylketone (FMK) and feruloylaldehyde (FAL). The presence of GSTP1-1 significantly accelerated the initial rate of GSH-mediated consumption of curcumin in 10 mM potassium phosphate, pH 7.0 and 1 mM GSH. GSTP1-1 kinetics determined using HPLC indicated substrate inhibition (apparent Km for curcumin of 25 ± 11 ÌM, and apparent Ki for curcumin of 8 ± 3 ÌM). GSTP1-1 was also shown to catalyze the reverse reaction, leading to the formation of curcumin from GSH adducts of FMK and FAL. Thus the effects of curcumin on the GSH-GST system may explain the chemopreventive activities assigned to it. µ1S. Suppression of inflammation by curcumin: Inflammation has been implicated in carcinogenesis. Numerous lines of evidence, both in vitro and in vivo, suggest that curcumin is a potent anti-inflammatory agent (47, 95, 113, 114, 128, 162, 225-229). Joe et al. (229) examined the effect of curcumin on arachidonic acid metabolism and secretion of lysosomal enzymes by rat peritoneal macrophages. They found that 10 ÌM curcumin treatment for 1 hour inhibited the incorpo-ration of arachidonic acid into the membrane lipids by 82%: prostaglandin E2 by 45%; leukotriene B4 by 61% and leukotriene C4 by 34%, respectively, but did not affect the release of arachidonic acid from macrophages stimulated by PMA. However, the secretion of 6-keto PGF1a was enhanced by 40% from macrophages preincubated with 10 ÌM curcumin. Curcumin also inhibited the secretion of colla-genase, elastase and hyaluronidase. These results thus showed that curcumin can control the release of inflammatory mediators such as eicosanoids and hydrolytic enzymes secreted by macrophages, thus explaining its anti-inflamma-tory properties. Reddy et al. examined the anti-inflammatory activity of curcumin on carrageenin-induced inflammation in rats (228). They found that curcumin, when given by gavage, lowered carrageenin-induced edema in the foot pads of rats. Interestingly. supplementation of diets with 1% curcumin (w/w) did not affect the inflammatory responses of animals to carrageenin injection. Yamamoto et al. examined the effect of curcumin on mammalian phospholipase D (PLD), phospha-tidylinositol- specific PLC and PLA2 from mouse macro-phage-like cell line J774.1 cells, sphingomyelinase from bovine brain and phosphatidylcholine-phospholipase C from Bacillus cereus (162). Curcumin inhibited several types of phospholipases, most effectively PLD among those tested. It also inhibited PMA-induced PLD activation in intact J774.1 cells. With respect to inflammation, curcumin has been found to inhibit the activation of free radical-activated transcription factors in vitro, such as NF-κ µ and AP-1, and to reduce the production of pro-inflammatory cytokines such as TNF, IL-1ß, and IL-8. The response to curcumin of inducible nitric oxide synthase (iNOS), an inflammationinduced enzyme that catalyzes the production of nitric oxide (NO) and a molecule that may lead to carcinogenesis, was studied by Chan et al., in vivo. (128). They found that in ex vivo cultured BALB/c mouse peritoneal macrophages, 1-20 ÌM curcumin reduced the production of iNOS mRNA in a concentration-dependent manner. Furthermore, they demonstrated that two oral treatments of 0.5 mL of a 10 ÌM solution of curcumin (92 ng/g of body weight) reduced iNOS mRNA expression in the livers of LPS-injected mice by 50-70%. Although many hold that curcumin needs to be given at dosages that are unattainable through diet to produce an in vivo effect, these investigators obtained potency at concentrations of nanomoles per gram of body weight. This efficacy was associated with two modifications in the preparation and feeding regimen : 1) an aqueous solution of curcumin was prepared by initially dissolving the compound in 0.5 N NaOH and then immediately diluting it in PBS; and 2) mice were fed curcumin at dusk after fasting. Inhibition was not observed in mice that were fed ad libitum, suggesting that food intake may interfere with the absorption of curcumin. B1T. Effect of curcumin on mutagenic effects of drugs: Numerous studies suggest that curcumin is a potent antimutagenic agent (230-247) that may contribute to its chemopreventive effects. Nagabhushan et al. examined the effect of curcumin on the mutagenicity of several envi-ronmental mutagens in the Salmonella microsome test with or without Aroclor 1254-induced rat-liver homogenate (S-9 mix) (230). With Salmonella typhimurium strain TA98 in the presence of S-9 mix, curcumin inhibited the mutagenicity of bidi (a rural cigarette where tobacco is wrapped in a dried leaf) and cigarette smoke condensates, tobacco and masheri extracts, benzo[a]pyrene and dimethyl benzo[a]anthracene. curcumin did not influence the mutagenicity without the S-9 mix of sodium azide, monoacetyl hydrazine and streptozocin in strain TA100 nor of 4nitrophenylenediamine in strain TA98. These observations indicated that curcumin may alter the metabolic activation and detoxification of mutagens. Shalini et al. examined the effect of curcumin on lipid peroxide-induced DNA damage (231). Curcumin was very effective in protection of DNA against peroxidative injury. Shalini et al. also investigated the effect of curcumin on the fuel smoke condensate (FSC)-induced DNA damage in human lymphocytes using fluorescence analysis of DNA unwinding (232). Curcumin protected DNA against FSC and PMA. Donatus et al. examined the effect of curcumin on paracetamol-induced cytotoxicity, lipid peroxidation, and glutathione depletion in rat hepatocytes (233). Paracetamol was selected as a model toxin because it is known to be bioactivated by 3-methylcholanthrene-inducible cytochrome P450, presumably to N-acetyl-p-benzoquinone imine, a reactive metabolite that upon overdosage causes protein and non-protein thioldepletion, lipid peroxidation and cytoto-xicity, the last measured as LDH leakage. At low con-centrations, curcumin protected hepatocytes significantly against paracetamol-induced lipid peroxidation, without protecting them against paracetamol-induced LDH leakage or paracetamol-induced GSH depletion. At 100 times the low-dose concentration of curcumin, protection against on lipid peroxidation was accompanied by a tendency to increase cellular GSH depletion and LDH leakage. It was concluded that curcumin’s cytoprotective effects at low dose and cytotoxic effects at high dose may be explained by a strong anti-oxidant capacity of curcumin and the ability of curcumin to conjugate with GSH. Abraham et al. investigated the effect of curcumin against Á-radiation-induced in vivo chromosomal damage using the mouse bone marrow micronucleus test (236). Oral administration of curcumin (5, 10, and 20 mg/kg b.w.) to mice significantly reduced the frequency of micronucleated polychromatic erythrocytes induced by whole body exposure to Á-radiation (L 15 Gy; 0.05 Gy/s). This effect was observed when curcumin was given after a single administration either 2 hours before or immediately after irradiation. The protective effects were observed in bone marrow cells sampled 24, 30 and 48 hours after exposure to radiation. Soudamini et al. used S. typhimurium strains TA 100 and TA 1535 to examine the mutagenicity and anti-mutagenicity of curcumin (237). Turmeric extract was found to inhibit microsomal activation-dependent mutagenicity of 2-aceta-midofluorene. Similar results were also obtained using curcumin. Oda et al. reported inhibitory effects of curcumin on SOS functions induced by UV irradiation in S. typhimurium TA 1535/pSK1002 and E. coli K-12 strain (238). Induction of the SOS gene (umuC) expression was assayed by measuring accumulated ß-galactosidase activity. They found that curcumin blocked umuC induction promoted by UV irradiation. In regard to another SOS response, the Weigle reactivation, they observed that curcumin effectively inhibited phage reactivation by UV irradiation. They also showed that mutagenesis induced by UV irradiation was suppressed by the addition of curcumin. Recently, Grandhean-Laquerriere et al. have studied the modulatory effect of curcumin on the UVB-induced cytokine expression by keratinocytes (248). Following UVB radiation treatment, expression of TNF-•, IL-6 and IL-8 by NCTC 2544 keratinocyte cell line was significantly enhanced both at the mRNA and protein level. The UVB also increased the IL-10 steady-state mRNAs level. Radiation-induced cytokine overexpression was accompanied by NF-κ B and AP-1 trans-cription factors. To investigate in keratinocytes the relative contributions of those transcription factors on UVB-mediated cytokine induction, cell cultures were supplemented with curcumin. Curcumin significantly inhibited NF-κ B activation by UVB, but AP-1 activation was unaffected. In parallel, curcumin decreased, in a dose-dependent manner, the UVB-mediated overexpression of all three pro-inflammatory cytokines and only exhibited a moderate enhancing influence on IL-10 expression. Deshpande et al. (239) investigated the effects of curcumin on the formation of benzo[a]pyrene-derived DNA adducts in vitro employing mouse liver S9. A dose-dependent decrease in binding of [3H]B(a)P metabolites to calf thymus DNA was observed in the presence of curcumins. Further studies employing mouse liver microsomes and individual compo-nents of curcumins, i.e. curcumin (C), demethoxycurcumin (dmC) and bisdemethoxycurcumin (bdmC), showed that all three components brought about dose-dependent inhibition of [3H]B(a)P-DNA adduct formation. Inhibitory activity was on the order C > dmC > bdmC. Investigations on the inhibitory effect of curcumin showed a dose-dependent decrease in cytochrome P450 and aryl hydrocarbon hydro-xylase (AhH) activity, resulting in increased unmetabolized B(a)P in the presence of curcumin. A comparison of the structures of curcumins with their activity profile suggested the importance of both parahydroxy (p-OH) and methoxy groups (-OCH3) in the structure-activity relationship. Removal of curcumin restorated cytochrome P450 activity and the levels of [3H]-B(a)P-DNA adducts. Thus these studies demonstrated that curcumin is effective in inhibiting [3H]B(a)P-derived DNA adducts by interfering with the metabolic enzymes and that its physical presence is essential for this effect. Anto et al. investigated the antimutagenic and anticarci-nogenic activity of natural and synthetic curcuminoids (240). They used five synthetic curcuminoids and three natural curcuminoids. The natural curcuminoids, curcumin I (diferu-loylmethane), curcumin ππ (feruloyl-p-hydroxycinnamoylme-thane) and curcumin III (bis-(phydroxycinnamoyl) methane), isolated from C longa were found to be potent inhibitors of mutagenesis and crotan oil-induced tumor promotion. Curcumin III produced 87% inhibition of 2-acetami-dofluorene (2-AAF)-induced mutagenesis, at a concentration of 100 Ìg/plate; curcumin ππ and curcumin I produced 70% and 68% inhibition at the same concentration. All the synthetic curcuminoids inhibited 2-AAF-induced mutage-nicity; salicyl- and anisyl-curcuminoids were the most active. Curcumin III most effectively antagonized the tumor promotor among the natural curcuminoids: while 90% of control animals had papillomas on the 10th week of tumor initiation, only 10% of the curcumin Iππ-treated animals, 20% of the curcumin ππ-treated animals and 40% of the curcumin I-treated animals had papillomas at that time. Salicylcurcu-minoid, which completely blocked papilloma formation in the 10th week, was the most potent anti-carcinogen among the synthetic curcuminoids. Piperonal curcuminoid also exhibited antipromotional activity. Soni and his coworkers examined the protective effect of curcumin on aflatoxin-induced mutagenicity and hepato-carcinogenicity using Salmonella tester strains TA 98 and TA 100 (241). Curcumin inhibited mutation frequency by more than 80% at concentrations of 2 Ìg/plate. Dietary admi-nistration of curcumin to rats significantly reduced the number of aflatoxin-induced Á-glutamyl transpeptidase-positive foci, which are considered precursors of hepato-cellular neoplasms. These results indicate the usefulness of curcumin in ameliorating aflatoxin-induced mutagenicity and carcinogenicity. Interestingly, Ahsan et al. showed that in the presence of Cu(II), curcumin caused breakage of calf thymus and supercoiled plasmid pBR322 DNA (242). The products were relaxed circles with no detectable linear forms. The metal ions tested, Mg(II), Ca(II), Fe(II) and Ni(II) were ineffective or less effective in the DNA breakage reaction. Cu(I) was shown to be an essential intermediate by using the Cu(I)-specific sequestering reagent neocuproine. The involvement of reactive oxygen species, such as H2O2 and 1O2 was established by the inhibition of DNA breakage by catalase and azide. Curcumin is also able to directly produce O2- and H2O2, and in the presence of Cu(II), OH is generated. Absorption spectra of curcumin in the presence of DNA indicated that a complex is formed between the two. Similarily, Antunes et al. examined the effects of curcumin on chromosomal damage induced by doxorubicin (DXR) in Chinese hamster ovary (CHO) cells (243). The DXR is a known free radical generator. Cells were treated with three concentrations of curcumin (2.5, 5, or 10 Ìg/ml), and then treated with DXR (1 Ìg/ml) during different phases of the cell cycle. Curcumin induced chromosomal damage in CHO cells, and treatment with curcumin did not reduce the clastogenicity of DXR. Instead, a statistically significant increase in the frequency of chromosomal damage was observed when curcumin was combined with DXR during the G1/S- S- and S/G2- phases of the cell cycle. The results clearly indicated the potentiating effect of curcumin on DXR-induced chromosomal damage. Kelly et al. examined the effect of curcumin on oxidative damage to cellular DNA (244). They pretreated Jurkat T-lymphocytes for 30 minutes with 0-25 ÌM curcumin and then challenged them with 25 ÌM H2O2. Afterwards, the cells were subjected to alkaline microgel electrophoresis (i.e. comet assay) to assess the extent of single-strand breaks in DNA. Curcumin alone actually induced DNA damage. This effect did not appear to reflect the DNA fragmentation associated with apoptosis because there was no proteolytic cleavage of poly-(ADP-ribose)-polymerase, an early marker of apoptosis. Curcumin-induced damage to DNA was prevented by pre-treatment of the cells with the lipophilic antioxidant •-tocopherol, suggesting that curcumin damaged DNA through oxygen radicals. Therefore, it was concluded that curcumin has prooxidant activity in cultured cells based on their opposite effects on DNA. Araujo et al. investigated the protective effect of thiourea, a hydroxyl-radical scavenger, on curcumin-induced chromo-somal aberrations in an in vitro mammalian cell system (245). Although known to scavenge free radicals and to inhibit clastogenesis in mammalian cells, curcumin has also been reported to induce a significant increase in the frequency of chromosomal aberrations in CHO cells. To investigate whether the clastogenic activity of curcumin in CHO cells in culture can be ascribed to a pro-oxidant behavior, mediated by free radical generation, experiments were carried out with the combination of curcumin (15 Ìg/ml) and thiourea (10, 20, or 40 Ìg/ml), a potent hydroxyl radical scavenger. The results showed that the clastogenic action of curcumin was stati-stically significantly decreased in the presence of thiourea. These data show that curcumin-induced chromosomal damage in CHO cells can be mediated by hydroxyl radical generation. How curcumin induces arrest in the G2/M-phase of the cell cycle is not well understood. It is possible that growth arrest is associated with structural changes in cellular organization during mitosis. Holy et al. found that curcumin disrupts the mitotic spindle structure and induces micronucleation in MCF-7 breast cancer cells (246). Treatment of MCF-7 breast cancer cells with 10-20 ÌM curcumin for 24-48 hÔurs arrested them in M-phase and inhibited DNA synthesis almost completely. Remarkably, arrested mitotic cells exhibited monopolar spindles and chromosomes did not undergo normal anaphase movements. After 48 hÔurs, most cells eventually left M-phase, and many formed multiple micronuclei instead of individual daughter nuclei. These observations indicate that the curcumin-induced G2/M arrest previously described for MCF-7 cells is due to the assembly of aberrant, monopolar mitotic spindles that are impaired in their ability to segregate chromosomes. The production of cells with extensive micronucleation after curcumin treatment suggests that at least some of the cytostatic effects of curcumin are due to its ability to disrupt normal mitosis, and raises the possibility that curcumin may promote genetic instability under some circumstances. Shukla et al. examined the antimutagenic potential of curcumin on chromosomal aberrations using in vivo chromosomal aberration assay in Wistar rats (247). Cyclophosphamide (CP), a well-known mutagen, was given by intraperitoneal (i.p.) injection at the dose of 40 mg/kg body weight (b.w.). Curcumin was given at the dose of 100 and 200 mg/kg b.w. through gastric intubation for seven consecutive days prior to CP treatment. The animals were sacrificed at the sampling time of 24 hÔurs after treatment, and their bone marrow tissue was analyzed for chromosomal damage and mitotic index. In CP-treated animals, a significant induction of chromosomal aberration was recorded with decrease in mitotic index. However, in curcumin-supplemented animals, no significant induction in chromosomal damage or change in mitotic index was recorded. In different curcumin-supple-mented groups, a dose-dependent significant decrease in CP induced clastogenicity was recorded. The incidence of aberrant cells was reduced by both doses of curcumin when compared to the CP-treated group. The anticytotoxic potential of curcumin towards CP was also evident as the mitotic index showed an increment. The study revealed the antigenotoxic potential of curcumin against CP -induced chromosomal mutations. These anti-mutagenic effects contribute to the chemopreventive potential of curcumin.
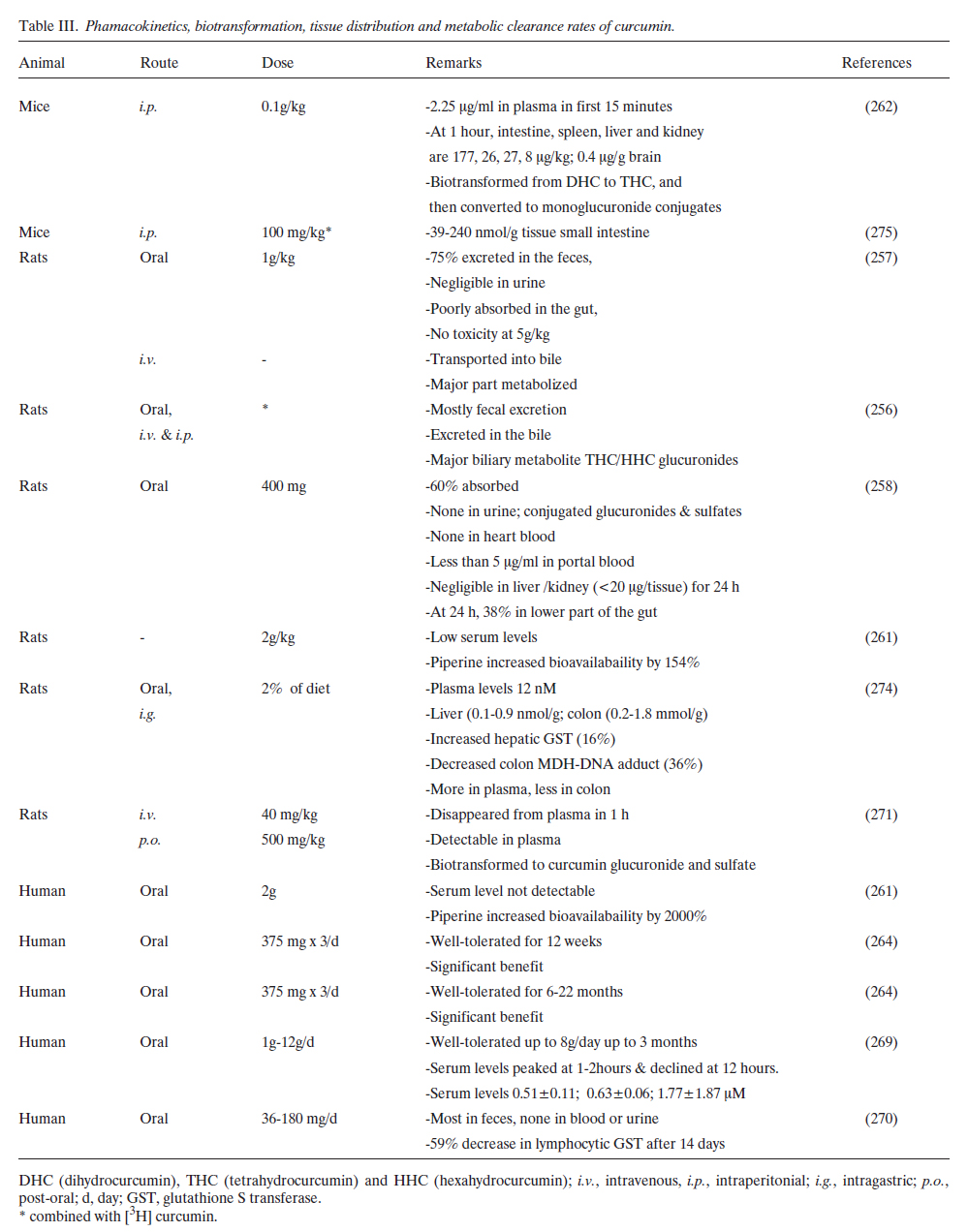
B1U. Curcumin can induce radioprotection: Several studies suggest that curcumin is radioprotective. Thresiamma et al. showed curcumin protects rats from radiation- induced toxicity (249). Whole body irradiation of rats (10 Gy as five fractions) produced lung fibrosis within 2 months, as assessed by increased lung collagen hydroxyproline and histo-pathology. Oral administration of curcumin (200 Ìmole/kg body weight) significantly reduced hydroxyproline. In serum and liver, irradiation-induced increases in lipid peroxidation were reduced significantly by curcumin treatment. The liver SOD and GSH peroxidase activity increases were also reduced significantly by curcumin, as was the frequency of micronucleated polychromatic erythrocytes in mice. In another study, Thresiamma et al. later investigated the protective effect of curcumin on radiation-induced geno-toxicity (250). They showed that induction of micronuclei and chromosomal aberrations produced by whole-body exposure of Á-radiation (1.5-3.0 Gy) in mice was significantly inhibited by oral administration of curcumin (400 Ìmoles/kg body weight), inhibited micronucleated polychromatic and normo-chromatic erythrocytes, significantly reduced the number of bone marrow cells with chromosomal aberrations and chromosomal fragments, and inhibited the DNA strand breaks produced in rat lymphocytes upon radiation as seen from the DNA unwinding studies. In contrast to these studies, Araujo et al. showed potentiation by curcumin of Á-radiation – induced chromosome aberrations in CHO cells (251). They treated the cells with curcumin (2.5, 5, and 10 Ìg/ml), and then irradiated (2.5 Gy) them during different phases of the cell cycle. curcumin at 10 Ìg/ml increased the chromosomal damage frequency. Curcumin did not have a protective effect against the clastogenicity of Á-radiation. Instead, an obvious increase in the frequencies of chromosome aberrations occurred when curcumin was given at 10 Ìg/ml plus Á-radiation during S- and G2/S- phases of the cell cycle. Why there is a difference in the results of Thresiamma et al. and Araujo et al. is unclear. Inano and Onoda investigated the radioprotective action of curcumin on the formation of urinary 8-hydroxy-2′-deoxy-guanosine (8-OHdG), tumorigenesis and mortality induced by Á-irradiation. The rats were examined for mammary and pituitary tumors for 1 year. Virgin rats received whole-body irradiation with 9.6 Gy and were fed a diet containing curcumin for 3 days before and/or 3 days after irradiation. After irradiation, all rats were assessed for 8-OHdG levels and survival was assessed daily for 30 days. The invstigators found that, in virgin rats irradiated with 3 Gy, the creati-nine-corrected concentration and total amount of 8-OHdG in the 24-hours urine samples were higher (~1.3-fold) than in the non-irradiated controls. Adding curcumin to the diet for 3 days before and/or 2 days after irradiation reduced the elevated 8-OHdG levels by 50-70%. Dietary curcumin (1%, w/w) significantly decreased the incidence of mammary and pituitary tumors. However, curcumin did not improve survival when administered for 3 days before and/or 3 days after irradiation (9.6 Gy). These findings demonstrate that curcumin can be used as an effective radioprotective agent to inhibit acute and chronic effects, but not mortality, after irradiation. How curcumin provides radioprotection is not fully understood. A few studies, however, indicate that curcumin inhibits the radiation-induced damage of specific proteins. Kapoor and Priyadarsini examined the free radical reactions of lysozyme (Lz), tryptophan and disulfides to curcumin in aqueous solution using a pulse radiolysis technique (252). The binding of curcumin with lysozyme was confirmed using absorption, fluorescence and stopped-flow techniques. The free radicals of curcumin generated after repairing radicals of disulfides, lysozyme and tryptophan absorbed meximally at 500-510 nm. This interaction may play a role in the anti-oxidant behavior of curcumin in protecting proteins. In contrast, Varadkar et al. examined the effect of curcumin on radiation-induced PKC activity isolated from the liver cytosol and the particulate fraction of unirradiated mice and mice irradiated at 5 Gy (253). Following irradiation, the PKC activity was increased in both cytosolic and particulate fractions. Curcumin inhibited the activated cytosolic and particulate PKC at very low concentrations. Since activation of PKC is one of the means conferring radioresistance on a tumor cell, suppression of PKC activity by curcumin might prevent the development of radioresistance. Another potential mechanism of radioprotection involves suppression of radiation-induced gene expression. Oguro and Yoshida examined the effect of curcumin on UVA-induced ornithine decarboxylase (ODC) and metallothionein (MT) gene expression in mouse skin (254). They showed that UVA induced MT mRNA in mouse skin; and 1,4-diazabicylo-[2,2,2] -octane (DABCO), a singlet oxygen scavenger, reduced UVA-mediated induction of MT mRNA (by 40%). UVA slightly enhanced TPA-mediated ODC mRNA induction, while it enhanced ODC enzyme activity by 70%. UVA additively intensified TPA-mediated MT mRNA induction. Curcumin dramatically inhibited both TPA- and TPA + UVA-induced expression of ODC and MT genes. B1V. Curcumin inhibits telomerase activity: Telomerase activity is linked to cellular proliferation, and its activation seems to be a mandatory step in carcinogenesis. Ramachandran et al. analyzed the effect of curcumin on telomerase activity in human mammary epithelial (MCF-10A) and breast cancer (MCF-7) cells (255). Telomerase activity in MCF-7 cells was 6.9-fold higher than that of human mammary epithelial cells. In MCF-7 cells, telomerase activity decreased with increasing concentrations of curcumin, inhibiting about 93.4% activity at 100 ÌM concentration. The inhibition of telomerase activity in MCF-7 cells may be due to down-regulation of hTERT expression. Increasing concentrations of curcumin caused a steady decrease in the level of hTERT mRNA in MCF-7 cells, whereas the level of c-myc mRNAs remained the same. These results suggest that curcumin inhibits telomerase activity by down-regulating hTERT expression in breast cancer cells and this down-regulation is not through the c-myc pathway. Since hTERT is known to be regulated by NF-κ B, it is possible that the down-regulation of hTERT is mediated through suppression of NF-κ B by curcumin. From the studies described above it is clear that curcumin interferes with multiple signal transduction pathways (see Figure 4), which could play a role in tumorigenesis. The suppression of these pathways may explain the antitumor effects of curcumin.
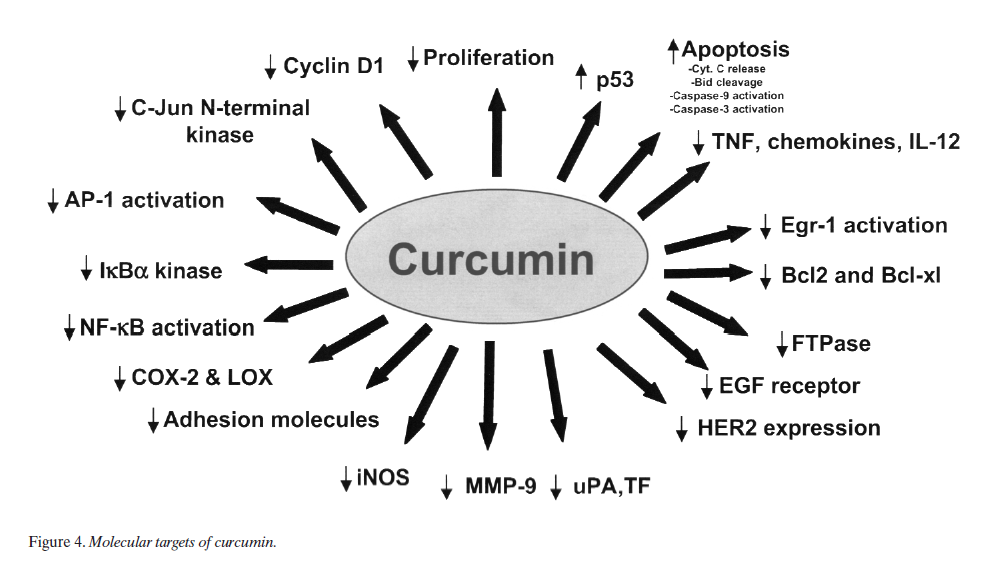
In vivo animal studies with curcumin
µ2A. Metabolism, pharmacokinetics, tissue distribution and clearance of curcumin: B2A1: Animal studies: Curcumin has been administered to the animals via a variety of routes including topical, systemic, intravenous and oral routes of administration (Table πππ) (5, 61, 224, 256-274). Wahlstrom et al. examined the uptake, distribution and excretion of curcumin administered orally to Sprague-Dawley rats (257). When administered at a dose of 1g/kg, about75% of curcumin was excreted in the feces, while negligible amounts of curcumin appeared in the urine. Measurements of blood plasma levels and biliary excretion showed that curcumin was poorly absorbed from the gut. No apparent toxic effects were seen after doses of up to 5 g/kg. When intravenously injected or when added to the perfusate of the isolated liver, curcumin was actively transported into bile, against con-centration gradients of several hundred times. The major part of the drug was, however, metabolized. In suspensions of isolated hepatocytes or liver microsomes 90% of the added curcumin was metabolized within 30 minutes. Holder et al. used curcumin labeled with deuterium and tritium to examine the pharmacokinetics (256). Oral and intraperitoneal doses of [3H] curcumin led to the fecal excretion of most of the radioactivity. When administered via either the intravenous or intraperitoneal route, [3H] curcumin was excreted in the bile of cannulated rats. Chemical ionization mass spectro-metry demonstrated that the major biliary metabolites were glucuronides of tetrahydrocurcumin and hexahydrocurcumin. A minor biliary metabolite was dihydroferulic acid together with traces of ferulic acid. Ravindranath et al. examined the absorption and tissue distribution of curcumin (400 mg) after oral administration in rats (258). About 60% of the dose was absorbed; no curcumin was detectable in urine. The urinary excretion of conjugated glucuronides and sulfates significantly increased. No curcumin was present in heart blood. Only traces (less than 5 Ìg/ml) in portal blood and negligible quantities in liver and kidney (< 20 Ìg/ tissue) were observed from 15 minutes up to 24 hours after administration of curcumin. At the end of 24 hours the concentration of curcumin remaining in the lower part of the gut, namely the cecum and large intestine, amounted to 38% of the quantity administered. The same investigators also examined the in vitro intestinal absorption of curcumin in rats (259). When everted sacs of rat intestines were incubated with 50-750-Ìg of curcumin, 30-80% of the added curcumin disappeared from the mucosal side. No curcumin was detectable in the serosal fluid. Less than 3% of the added curcumin was found in the tissue at the highest concentration. In experiments with [3H] curcumin, 5-6% of added radioactivity was found in the serosal side. TLC examination of the mucosal extract showed the presence of 2 compounds, 1 corresponding to curcumin and the other to a less polar, colourless compound. The serosal fluid had no curcumin, but there was a compound whose RF was identical with the colourless compound present in the mucosal side. These experiments indicated that curcumin undergoes transformation during absorption from the intestine. Like Holder et al., Ravindranath examined the metabolism of curcumin using [3H] curcumin (256, 260). Radioactivity was detectable in blood, liver and kidney following doses of 400, 80, or 10 mg of [3H] curcumin. The major route of elimination of the label was the feces; urinary excretion was very low regardless of the dose. At the lower doses of 80 mg and 10mg of [3H] curcumin, mostofthe label was excreted within 72 hours, while with 400 mg, considerable amounts of the label was present in the tissues 12 days after dosage. The percentage of curcumin absorbed (60-66% of the given dose) remained constant regardless of the dose administered. The above mentioned studies indicate poor absorption, rapid metabolism and excretion of curcumin. Therefore, it is unlikely that substantial concentrations of curcumin occur in the body after ingestion. However, Pan et al. investigated the pharmacokinetic properties of curcumin in mice after i.p. administration (262). After a dose of 0.1 g/kg, about 2.25 Ìg/ml of curcumin appeared in the plasma in the first 15 minutes. One hour after administration, the levels of curcumin in the intestines, spleen, liver and kidneys were 177, 26, 27 and 8 Ìg/g, respectively. Only traces (0.4 Ìg/g) were observed in the brain at 1 hour. When plasma was examined for the nature of the metabolites of curcumin, by reverse-phase HPLC, two putative conjugates were observed. Treatment of the plasma with P-glucuronidase decreased the concentrations of these two putative conjugates and tetrahydrocurcumin (THC) and curcumin were detected. To investigate the nature of these glucuronide conjugates in vivo, the plasma was analyzed by electrospray. The chemical structures of these metabolites, determined by mass spectro-metry analysis, suggested that curcumin was first bio-transformed to dihydrocurcumin and THC and that these compounds subsequently were converted to monoglucuronide conjugates. Because THC is one of the major metabolites of curcumin, its stability at different pH values was examined. THC was very stable in 0.1 M phosphate buffers of various pH values. Moreover, THC was more stable than curcumin in 0.1 M phosphate buffer, pH 7.2. These results suggested that curcumin-glucuronoside, dihydrocurcumin-glucurono-side, THC-glucuronoside and THC are major metabolites of curcumin in vivo. Sharma et al. tested the hypothesis that 14 days of dietary curcumin (2%) affects biomarkers relevant to cancer chemoprevention in the rat (274). Levels of inducible COX2, as reflected by PGE2 production by blood leukocytes, were measured ex vivo. Total GST activity and adducts of malondialdehyde with DNA (M(1)G), which reflect endogenous lipid peroxidation, were measured in colon mucosa, liver and blood leukocytes. Curcumin and its metabolites were analyzed by HPLC in plasma and its pharmacokinetics were compared following a diet containing 2% curcumin versus intragastric (i.g.) administration of curcumin suspended in an amphiphilic solvent. The curcumin diet did not alter any of the markers in the blood but increased hepatic GST by 16% and decreased colon M(1)G levels by 36% when compared with controls. Administration of carbon tetrachloride during the treatment period increased colon M(1)G levels, and this increase was prevented by dietary curcumin. Dietary curcumin yielded low drug levels in the plasma, between 0 and 12 nM, whereas tissue con-centrations of curcumin in liver and colon mucosa were 0.1-0.9 nmol/g and 0.2-1.8 Ìmol/g, respectively. In comparison with dietary administration, suspended curcumin given i.g. resulted in more curcumin in the plasma but much less in the colon mucosa. The results show that curcumin mixed with the diet achieves drug levels in the colon and liver sufficient to explain the pharmacological activities observed and suggest that this mode of administration may be preferable for the chemoprevention of colon cancer. B2A2: Studies in humans: Animal studies have revealed that curcumin has poor bioavailability due to its rapid metabolism in the liver and intestinal wall. Therefore, Shoba et al. exa-mined the effect of combining piperine, a known inhibitor of hepatic and intestinal glucuronidation, on the bioavailability of curcumin in rats and healthy human volunteers (261). When curcumin was given alone, in the dose 2 g/kg to rats, moderate serum concentrations were achieved over a period of 4 hours. Concomitant administration of piperine 20 mg/kg increased the serum concentration of curcumin for a short period of 1 – 2 hours. The time to maximum concentration was significantly increased (p < 0.02), while elimination half-life and clearance significantly decreased (p < 0.02), and the bioavailability was increased by 154%. On the other hand in humans after a dose of 2 g curcumin alone, serum levels were either undetectable or very low. Concomitant administration of piperine 20 mg produced much higher concentrations from 0.25 to 1 hour (p < 0.01 at 0.25 and 0.5 hour; p < 0.001 at 1 hour), and the increase in bioavailability was 2000%. Investigators did not observe any adverse effect. The study showed that in the dosages used, piperine enhances the serum concentration, extent of absorption and bioavailability of curcumin in both rats and humans with no adverse effects. The systemic bioavailability of curcumin is low, so that its pharmacological activity may be mediated, in part, by curcumin metabolites. To investigate this possibility, Ireson et al. compared curcumin metabolism in human and rat hepatocytes in suspension with that in rats in vivo (271). Analysis by HPLC with detection at 420 and 280 nm permitted characterization of metabolites with intact diferoylmethane structure and increased saturation of the heptatrienone chain. Chromatographic inferences were corroborated by mass spectrometry. The major metabolites in suspensions of human and rat hepatocytes were identified as hexahydrocurcumin and hexahydrocurcuminol. In rats, in vivo, curcumin administered i.v. (40 mg/kg) disappeared from the plasma within 1 hour of dosing. After p.o. administration (500 mg/kg), the parent drug was present in plasma at levels near the detection limit. The major products of curcumin biotransformation identified in rat plasma were curcumin glucuronide and curcumin sulfate, whereas hexahydro-curcumin, hexahydrocurcuminol and hexahydrocurcumin glucuronide were present in small amounts. To test the hypothesis that curcumin metabolites resemble their pro-genitor in that they can inhibit COX2 expression, curcumin and four of its metabolites at a concentration of 20 ÌM were compared in terms of their ability to inhibit phorbol ester-induced PGE2 production in human colonic epithelial cells. Curcumin reduced PGE2 levels to preinduction levels, whereas tetrahydrocurcumin, previously shown to be a murine metabolite of curcumin, hexahydrocurcumin and curcumin sulfate, had only weak PGE2 inhibitory activity and hexahydrocurcuminol was inactive. The results suggest that (a) the major products of curcumin biotransformation by hepatocytes occur only at low abundance in rat plasma after curcumin administration; and (b) metabolism of curcumin by reduction or conjugation generates species with reduced ability to inhibit COX2 expression. Because the gastrointe-stinal tract seems to be exposed more prominently to unmetabolized curcumin than any other tissue, the results supported the clinical evaluation of curcumin as a colorectal cancer chemopreventive agent. Indirect evidence suggests that curcumin is metabolized in the intestinal tract. To investigate this notion further, Ireson et al. explored curcumin metabolism in subcellular fractions of human and rat intestinal tissue, compared it with metabolism in the corresponding hepatic fractions and studied curcumin metabolism in situ in intact rat intestinal sacs (272). Curcumin glucuronide was identified in intestinal and hepatic microsomes and curcumin sulfate, tetrahydro-curcumin and hexahydrocurcumin were found as curcumin metabolites in intestinal and hepatic cytosol from humans and rats. The extent of curcumin conjugation was much greater in intestinal fractions from humans than in those from rats, whereas curcumin conjugation was less extensive in hepatic fractions from humans than in those from rats. The curcumin-reducing ability of cytosol from human intestinal and liver tissue exceeded that observed with the corresponding rat tissue by factors of 18 and 5, respectively. Curcumin sulfate was identified in incubations of curcumin with intact rat gut sacs. Curcumin was sulfated by human phenol sulfotransferase isoenzymes SULT1A1 and SULT1A3.

Equine alcohol dehydrogenase catalyzed the reduction of curcumin to hexahydrocurcumin. The results show that curcumin undergoes extensive metabolic conjugation and reduction in the gastrointestinal tract and that there is more metabolism in human than in rat intestinal tissue. The pharmacological implications of the intestinal metabolism of curcumin should be taken into account in the design of future chemoprevention trials of this dietary constituent. B2B. Anticarcinogenic effects of curcumin: Several studies indicate that curcumin is a potent chemopreventive agent (137, 151, 232, 274-329). It can suppress both tumor initiation and tumor promotion (see Table IV). The initiation of tumors induced by benzo [a] pyrene, 7, 12 dimethylbenz [a] anthracene (DMBA), 12-O-tetradecanoylphorbol-13-acetate, azoxymethane, 1, 2 dimethylhydrazine, dimethylnitrosamine, 2-amino 1-methyl-6-phenylimidazo [4,5-b]pyridine (PhIP; a human dietary carcinogen), 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK), Á-radiation and other carcinogens and tumor promotion induced by PMA have been shown to be suppressed by curcumin (137, 278-281, 284-288, 290, 293, 294, 296, 297, 299, 300, 304, 305, 307, 308, 311, 314, 316, 320, 321, 328). Curcumin is also known to reverse aflatoxin-induced liver damage and prevent fuel smoke condensate-induced DNA damage (232, 280). The tumorigenesis of the skin, mammary gland, oral cavity, forestomach, oesophagus, stomach, intestine, colon, lung, and liver have been shown to be suppressed by curcumin (137, 274, 277, 278, 282, 284-289, 292, 294-296, 299, 303-305, 307, 313, 316, 320, 328). To explain the anticarcinogenic effects of curcumin on different tumors, a wide variety of mechanisms have been implicated, including inhibition of ROI, suppression of inflammation, down-regulation of ODC, inhibition of cell proliferation, inhibition of cytochrome P450 isozymes, induction of GSH, suppression of certain oncogenes (e.g; cHa-ras, c-jun and c-fos), inhibition of transcription factors NF-κ B and AP-1, suppression of COX2, inhibition of cell cycle-related proteins (PCNA, cyclin E, p34 cdc2), inhibition of chromosomal damage, inhibition of oxidation of DNA bases, inhibition of malondialdehyde DNA adduct formation, inhibition of tumor implantation, inhibition of protein tyrosine kinase and protein kinase C activity, inhibition of biotransformation of carcinogens and induction of GST activity (137, 232, 274, 282, 286-288, 290, 295, 296, 303, 306, 308, 314, 322, 323). Curcumin has also been shown to suppress the replication of viruses (such as EBV) which contribute to tumorigenesis (327). Although in most studies curcumin was administered in the diet, some studies employed topical application to tumors, especially studies of skin tumorigenesis (137, 277, 278, 284, 287, 296, 297). Up to 2% curcumin has been used in the diet (274, 306). The chemopreventive activity of curcumin was observed when it was administered prior to, during and after carcinogen treatment as well as when it was given only during the promotion/progression phase (starting late in prema-lignant stage) of colon carcinogenesis (306).

Hecht et al. investigated the chemopreventive effect of curcumin against lung tumor induction in A/J mice by the tobacco smoke carcinogens BaP and NNK (311). Treatment with dietary curcumin (2000 ppm) from 1 week after carcinogen treatment until termination had no effect on lung tumor multiplicity. A dose of 500 ppm curcumin could prevent rat tongue carcinogenesis (288). There are some reports about the organ specificity of the chemopreventive effects of curcumin (294). Dietary curcumin had little or no effect on NNK-induced lung carcinogenesis and DMBA-induced breast carcinogenesis in mice. Poor circulating bioavailability of curcumin may account for the lack of lung and breast carcinogenesis inhibition (297). Collett et al. investigated the effects of curcumin on apoptosis and tumorigenesis in male Apc(min) mice treated with the human dietary carcinogen PhIP (326). The intestinal epithelial apoptotic index in response to PhIP treatment was approximately twice as great in the wild-type C57BL/6 APC(+/+) strain as in Apc(min) mice. PhIP promoted tumor formation in Apc(min) proximal small intestine. Curcumin enhanced PhIP-induced apoptosis and inhibited PhIP-induced turnorigenesis in the proximal small intestine of Apc(min) mice. Mahmoud et al. investigated the effect of curcumin for the prevention of tumors in C57BL/6J-Min/+ (Min/+) mice which bear germline mutation in the Apc gene and spontaneously develop numerous intestinal adenomas by 15 weeks of age (332). At a dietary level of 0.15 %, curcumin decreased tumor formation in Min/+ mice by 63%. Examination of intestinal tissue from the treated animals showed that tumor prevention by curcumin was associated with increased enterocyte apoptosis and proliferation. Curcumin also decreased expression of the oncoprotein beta-catenin in the enterocytes of the Min/+ mouse, an observation previously associated with an antitumor effect. Recently, Perkins et al. also examined the preventive effect of curcumin on the development of adenomas in the intestinal tract of the C57B1/6J Min/+ mouse, a model of human familial APC (275). These investigators explored the link between its chemopreventive potency in the Min/+ mouse and levels of drug and metabolites in target tissue and plasma. Mice received dietary curcumin for 15 weeks, after which adenomas were enumerated. Levels of curcumin and metabolites were determined by high-performance liquid chromatography in plasma, tissues and feces of mice after either long-term ingestion of dietary curcumin or a single dose of [14C] curcumin (100 mg/kg) via the i.p. route. Whereas curcumin at 0.1 % in the diet was without effect, at 0.2 and 0.5%, it reduced adenoma multiplicity by 39 and 40%, respectively, compared with untreated mice. Hematocrit values in untreated Min/+ mice were drastically reduced compared with those in wild-type C57BI/6J mice. Dietary curcumin partially restored the suppressed hematocrit. Traces of curcumin were detected in the plasma. Differences in its concentration in the small intestinal mucosa, between 39 and 240 nmol/g of tissue, reflected differences in dietary concentration. [14C]Curcumin disappeared rapidly from tissues and plasma within 2-8 hours after dosing. The group concluded that curcumin may be useful in the chemo-prevention of human intestinal malignancies related to Apc mutations. The comparison of dose, resulting curcumin levels in the intestinal tract, and chemopreventive potency suggested tentatively that a daily dose of 1.6 g of curcumin is required for efficacy in humans. A clear advantage of curcumin over nonsteroidal antiinflammatory drugs is its ability to decrease intestinal bleeding linked to adenoma maturation. B2C: Antitumor effects of curcumin in animals: While there are numerous studies on the prevention of cancer by curcumin, there is very little known about the therapeutic effects of curcumin. Busquets et al. showed that systemic administration of curcumin (20 Ìg/kg body weight) for 6 consecutive days to rats bearing the highly cachectic Yoshida AH-130 ascites hepatoma inhibited tumor growth (31 % of total cell number) (333). Dorai et al. examined the efficacy of curcumin in vivo for suppressing the growth of LNCaP cells as heterotopically implanted tumors in nude mice (334). The LNCaP prostate cancer cells were grown, mixed with Matrigel, and injected subcutaneously into nude mice that received a synthetic diet containing 2% curcumin for up to 6 weeks. At the end point, sections taken from the excised tumors were evaluated for pathology, cell proliferation, apoptosis and vascularity. The results showed that curcumin markedly decreases the extent of cell proliferation as measured by the BrdU incorporation assay and a significant increase in the extent of apoptosis as measured by an in situ cell death assay. Moreover, a significant decrease in the microvessel density as measured by the CD31 antigen staining was also seen. These results thus suggest that curcumin could be a therapeutic anti-cancer agent, as it significantly inhibits prostate cancer growth and has the potential to prevent the progression of this cancer to its hormone refractory state. Menon et al. examined the effect of curcumin on lung metastasis induced by B16F-10 melanoma cells in female C57BL/6 mice (335). Curcumin significantly inhibited lung tumor formation (89%) and significantly increased the life span (144%). Moreover, lung collagen hydroxyproline and serum sialic acid levels were found to be significantly lower in treated animals compared to the untreated controls. Curcumin treatment (10 Ìg/ml) significantly inhibited the invasion of B16F-10 melanoma cells across the collagen matrix of the Boyden chamber. Gelatin Zymographic analysis of a trypsin-activated B16F-10 melanoma cell sonicate revealed that curcumin did not have any metalloproteinase activity. Nor did curcumin treatment inhibit the motility of B16F-10 melanoma cells across a polycarbonate filter in vitro. These findings suggest that curcumin inhibits the invasion of B16F-10 melanoma cells by inhibition of metalloproteinases, thereby inhibiting lung metastasis.
Clinical Studies
C1. Phase I clinical trials: Sharma et al. investigated the pharmacodynamics and pharmacokinetics of curcumin in humans, a dose-escalation pilot study at doses between 440 and 2200 mg/day of Curcuma extracts, containing 36-180 mg of curcumin (270). Fifteen patients with advanced colorectal cancer refractory to standard chemotherapies received Curcuma extract daily for up to 4 months. The activity of GST and levels of a DNA adduct (M(1)G) formed by malondial-dehyde, a product of lipid peroxidation and prostaglandin biosynthesis, were measured in patients’ blood cells. Oral Curcuma extract was well tolerated and dose-limiting toxicity was not observed. Neither curcumin nor its metabolites were detected in blood or urine, but curcumin was recovered from feces. curcumin sulfate was identified in the feces of one patient. Ingestion of 440 mg of Curcuma extract for 29 days was accompanied by a 59% decrease in lymphocytic GST activity. At higher dose levels, this effect was not observed. Leukocytic M(l)G levels were constant within each patient and unaffected by treatment. Radiologically stable disease was demonstrated in five patients for 2-4 months of treatment. The results suggested that (a) Curcuma extract can be administered safely to patients at doses of up to 2.2 g daily, equivalent to 180 mg of curcumin; (b) curcumin has low oral bioavailability in humans and may undergo intestinal metabolism; and (c) larger clinical trials of Curcuma extract are merited. C2. Phase II clinical trials: Cheng et al. investigated the toxi-cology, pharmacokinetics and biologically effective dose of curcumin in humans (269). Their prospective phase – I/II study evaluated the effect of curcumin in patients with one of the following five high-risk conditions: 1) recently resected urinary bladder cancer; 2) arsenic Bowen’s disease of the skin; 3) uterine cervical intraepithelial neoplasm (CIN); 4) oral leucoplakia; and 5) intestinal metaplasia of the stomach. Curcumin was given orally for 3 months. Biopsy of the lesion sites was done immediately before and 3 months after starting curcumin treament. The starting dose was 500 mg/day. If no toxicity of grade ππ or higher was noted in at least 3 successive patients, the dose was then escalated to another level in the order of 1,000, 2,000, 4,000, 8,000 and 12,000 mg/day. The concentration of curcumin in serum and urine was determined by HPLC. A total of 25 patients were enrolled in this study. There was no treatment-related toxicity up to 8,000 mg/day. Beyond 8,000 mg/day, the bulky volume of the drug was unacceptable to the patients. The serum concentration of curcumin usually peaked at 1 to 2 hour after oral intake of crucumin and gradually declined over the next 12 hours. The average peak serum concentrations after taking 4,000 mg, 6,000 mg and 8,000 mg of curcumin were 0.51±0.11 ÌM, 0.63 ± 0.06 ÌM and 1.77 ± 1.87 ̪, respectively. Urinary excretion of curcumin was undetectable. One of 4 patients with CIN and 1 of 7 patients with oral leucoplakia proceeded to develop frank malignancies in spite of curcumin treatment. In contrast, histological improvement of precancerous lesions was seen in 1 of 2 patients with recently resected bladder cancer, 2 of 7 patients with oral leucoplakia, 1 of 6 patients with intestinal metaplasia of the stomach, 1 of 4 patients with CIN and 2 of 6 patients with Bowen’s disease. In conclusion, this study demonstrated that curcumin is not toxic to humans up to 8,000 mg/day when taken by mouth for 3 months. These results also suggested a biological effect of curcumin in the chemoprevention of cancer. Curcumin has shown wide-ranging chemopreventive activity in preclinical carcinogenic models, in which it inhibits COX2 at the transcriptional level. COX2 has been implicated in the development of many human cancers. Plummer et al. explored the inhibition of COX2 activity as a systemic biomarker of drug efficacy, a biomarker of potential use in clinical trials of many chemopreventive drugs known to inhibit this enzyme (268). They measured COX2 protein induction and PGE2 production in human blood after incubation with LPS. When 1 ÌM curcumin was added in vitro to blood from healthy volunteers, LPS-induced COX2 protein levels and concomitant PGE2 production were reduced by 24% and 41%, respectively. To test whether effects on COX2 activity could also be measured after oral dosing in humans, these workers conducted a dose escalation pilot study of a standardized formulation of Curcuma extract in 15 patients with advanced colorectal cancer. Basal and LPS-mediated PGE2 production was measured in blood, twice before treatment and on days 1, 2, 8 and 29 of treatment. Analysis of basal and LPS-induced PGE2 production during treatment demonstrated a trend toward dose-dependent inhibition ( p < 0.005 by regression analysis), but there was no significant difference compared with values from pretreatment time-points. Lal et al. administered curcumin orally to patients suffering from chronic anterior uveitis (CAU) at a dose of 375 mg three times a day for 12 weeks. Of 53 patients enrolled, 32 completed the 12-week study (264). They were divided into two groups: one group of 18 patients received curcumin alone, whereas the other group of 14 patients, who had a strong PPD reaction, in addition received antitubercular treatment. The patients in both the groups started improving after 2 weeks of treatment. All the patients who received curcumin alone improved, whereas the group receiving antitubercular therapy along with curcumin had a response rate of 86%. Follow-up of all the patients for the next 3 years indicated a recurrence rate of 55% in the first group and of 36% in the second group. Four of 18 (22%) patients in the first group and 3 of 14 patients (21%) in the second group lost their vision in the follow-up period due to various complications in the eyes, e.g. vitritis, macular edema, central venous block, cataract formation and glaucomatous optic nerve damage. None of the patients reported any side-effect of the drug. The efficacy of curcumin and recurrences following treatment are comparable to corticosteroid therapy, which is presently the only available standard treatment for this disease. The lack of side-effects with curcumin is its greatest advantage compared with cortico steroids. Curcumin has also been used for the treatment of patients suffering from idiopathic inflammatory orbital pseudotumors (267). curcumin was administered orally at a dose of 375 mg/3 times/day orally for a period of 6-22 months in eight patients. They were followed-up for a period of 2 years at 3-month intervals. Five patients completed the study, of which four recovered completely and one experienced some persistent limitation of movement but the swelling regressed completely. No side-effect was noted in any patient and there was no recurrence. The study suggests that curcumin could be used as a safe and effective drug in the treatment of idiopathic inflammatory orbital pseudotumors. C3. Epidemiological correlates of curcumin. It has been documented that cancer of the colon, breast, prostate and lung, while most common in the United States, is not as prevalent in countries such as India where curcumin is frequently consumed (336). Mohandas et al. examined the epidemiology of digestive tract cancers within India (263). They found a low incidence of large bowel cancers in Indians and attributed this to high intake of starch and the presence of natural antioxidants such as curcumin in Indian cooking. The role of hereditary factors has been evaluated in a few studies. Some studies have reported the occurrence of both FAP and HNPCC in India. Adenoma is rare in elderly Indians undergoing colonoscopy, even in those with large bowel cancers. They also reported that small bowel cancers are extremely rare in India. In conclusion, the incidence of large and small bowel adenomas and cancers is low in Indians. An increase in the incidence of large bowel cancers in immigrants and urban Indians compared to rural populations supports a role for environmental risk factors including diet.
Conclusion
From numerous studies as described above, it is quite apparent that curcumin has tremendous potential for prevention and therapy of various cancers. Several cellular proteins have been identified with which curcumin directly interacts (see Table V). One of the major problem with curcumin is its rapid clearance from the body. Whether rapid clearance is a disadvantage or an advantage is not clear at present. In spite of it, whether administered orally, i.p., or systemically, curcumin appears to have biological effects both in animals and humans. No natural agent has yet been described which modulates so many signal transduction pathways as curcumin does. Is that to our disadvantage? Probably not. Most cancer biologists are of the opinion that tumor cells use multiple pathways to escape host-defense mechanisms. Thus the drug which is specific for inhibition of one signal transduction pathway in the tumor cell may not be adequate. Finding a drug that attacks multiple pathways and yet is pharmacologically safe is difficult. Curcumin, however, is one such drug that meets this criterion. The inability to patent curcumin, because of its natural occurance and well-established, long history of dietary use, prevents the involvement of pharmaceutical industry to examine this drug closely as a therapeutic or preventive agent. The structure of curcumin can serve as lead compound for the design of a better molecule that is also patentable. It is our hope that other nonprofit organizations see the value in this compound and provide the kind of evidence needed to help cancer patients.
References
- Rao DS, Sekhara NC, Satyanarayana MN and Srinivasan M: Effect of curcumin on serum and liver cholesterol levels in the rat. J Nutr 1307-15, 1970.
- Patil TN and Srinivasan M: Hypocholesteremic effect of curcumin in induced hypercholesteremic rats. Indian J Exp Biol 9: 167-9, 1971.
- Keshavarz K: The influence of turmeric and curcumin on cholesterol concentration of eggs and tissues. Poult Sci 55: 1077-83, 1976.
- Soudamini KK, Unnikrishnan MC, Soni KB and Kuttan R: Inhibition of lipid peroxidation and cholesterol levels in mice by curcumin. Indian J Physiol Pharmacol 36: 239-43, 1992.
- Soni KB and Kuttan R: Effect of oral curcumin administration on serum peroxides and cholesterol levels in human volunteers. Indian J Physiol Pharmacol 36: 273-5, 1992.
- Hussain MS and Chandrasekhara N: Effect on curcumin on cholesterol gall-stone induction in mice. Indian J Med Res 96: 288-91, 1992.
- Asai A and Miyazawa T: Dietary curcuminoids prevent high-fat diet-induced lipid accumulation in rat liver and epididymal adipose tissue. J Nutr 131: 2932-5, 2001.
- Ramirez-Tortosa MC, Mesa MD, Aguilera MC, Quiles JL, Baro L, Ramirez- Tortosa CL, Martinez-Victoria E and Gil A: Oral admi-nistration of a turmeric extract inhibits LDL oxidation and has hypocholesterolemic effects in rabbits with experimental athe-rosclerosis. Atherosclerosis 147: 371-8, 1999.
- Naidu KA and Thippeswamy NB: Inhibition of human low density lipoprotein oxidation by active principles from spices. Mol Cell Biochem 229: 19-23, 2002.
- Patro BS, Rele S, Chintalwar GJ, Chattopadhyay S, Adhikari S and Mukherjee T: Protective activities of some phenolic 1,3-diketones against lipid peroxidation: possible involvement of the 1,3-diketone moiety. Chembiochem 3: 364-70, 2002.
- Srivastava R, Puri V, Srimal RC and Dhawan BN: Effect of curcumin on platelet aggregation and vascular prostacyclin synthesis. Arzneimittelforschung 36: 715-7, 1986.
- Srivastava KC, Bordia A and Verma SK: Curcumin, a major component of food spice turmeric (Curcuma longa) inhibits aggregation and alters eicosanoid metabolism in human blood platelets. Prostaglandins Leukot Essent Fatty Acids 52: 223-7, 1995.
- Srivastava R, Dikshit M, Srimal RC and Dhawan BN: Anti-thrombotic effect of curcumin. Thromb Res 40: 413-7, 1985.
- Dikshit M, Rastogi L, Shukla R and Srimal RC: Prevention of ischaemia-induced biochemical changes by curcumin & quinidine in the cat heart. Indian J Med Res 101: 31-5, 1995.
- Nirmala C and Puvanakrishnan R: Effect of curcumin on certain lysosomal hydrolases in isoproterenol-induced myocardial infarction in rats. Biochem Pharmacol 51: 47-51, 1996.
- Nirmala C and Puvanakrishnan R: Protective role of curcumin against isoproterenol induced myocardial infarction in rats. Mol Cell Biochem 159: 85-93, 1996.
- Venkatesan N: curcumin attenuation of acute adriamycin myocardial toxicity in rats. Br J Pharmacol 124: 425-7, 1998.
- Srinivasan M: Effect of curcumin on blood sugar as seen in a diabetic subject. Indian J Med Sci 26: 269-70, 1972.
- Babu PS and Srinivasan K: Influence of dietary curcumin and cholesterol on the progression of experimentally induced diabetes in albino rat. Mol Cell Biochem 152: 13-21, 1995.
- Babu PS and Srinivasan K: Hypolipidernic action of curcumin, the active principle of turmeric (Curcuma longa) in streptozotocin induced diabetic rats. Mol Cell Biochem 166: 169-75, 1997.
- Suresh Babu P and Srinivasan K: Amelioration of renal lesions associated with diabetes by dietary curcumin in streptozotocin diabetic rats. Mol Cell Biochem 181: 87-96, 1998.
- Arun N and Nalini N: Efficacy of turmeric on blood sugar and polyol pathway in diabetic albino rats. Plant Foods Hum Nutr 57: 41-52, 2002.
- Deodhar SD, Sethi R and Srimal RC: Preliminary study on antirheumatic activity of curcumin (diferuloyl methane). Indian J Med Res 71: 632-4, 1980.
- Natarajan C and Bright JJ: Curcumin inhibits experimental allergic encephalomyelitis by blocking IL-12 signaling through Janus kinase-STAT pathway in T lymphocytes. J Immunol 168: 6506-13, 2002.
- Lim GP, Chu T, Yang F, Beech W, Frautschy SA and Cole GM: The curry spice curcumin reduces oxidative damage and amyloid pathology in an Alzheimer transgenic mouse. J Neurosci 21: 8370-7, 2001.
- Frautschy SA, Hu W, Kim P, Miller SA, Chu T, Harris-White ME and Cole GM: Phenolic anti-inflammatory antioxidant reversal of a beta-induced cognitive deficits and neuropathology. Neurobiol Aging 22: 993-1005, 2001.
- Sui Z, Salto R, Li J, Craik C and Ortiz de Montellano PR: Inhibition of the HIV-1 and HIV-2 proteases by curcumin and curcumin boron complexes. Bioorg Med Chem 1: 415-22, 1993.
- Li CJ, Zhang LJ, Dezube BJ, Crumpacker CS and Pardee AB: Three inhibitors of type 1 human immunodeficiency virus long terminal repeat-directed gene expression and virus replication. Proc Natl Acad Sci USA 90: 1839-42, 1993.
- Jordan WC and Drew CR: Curcumin–a natural herb with anti-HIV activity. J Natl Med Assoc 88: 333, 1996.
- Mazumder A, Nearnati N, Sunder S, Schulz J, Pertz H, Eich E and Pommier Y: curcumin analogs with altered potencies against HIV-1 integrase as probes for biochemical mechanisms of drug action. J Med Chem 40: 3057-63, 1997.
- Barthelemy S, Vergnes L, Moynier M, Guyot D, Labidalle S and Bahraoui E: Curcumin and curcumin derivatives inhibit Tat-mediated transactivation of type 1 human immunodeficiency virus long terminal repeat. Res Virol 149: 43-52, 1998.
- Sidhu GS, Singh AK, Thaloor D, Banaudha KK, Patnaik GK, Srimal RC and Maheshwari RK: Enhancement of wound healing by curcumin in animals. Wound Repair Regen 6: 167-77, 1998.
- Phan TT, See P, Lee ST and Chan SY: Protective effects of curcumin against oxidative damage on skin cells in vitro: its implication for wound healing. J Trauma 51: 927-31, 2001.
- Shahed AR, Jones E and Shoskes D: Quercetin and curcumin up-regulate antioxidant gene expression in rat kidney after ureteral obstruction or ischemia/reperfusion injury. Transplant Proc 33: 2988, 2001.
- Morikawa T, Matsuda H, Ninomiya K and Yoshikawa M: Medi-cinal foodstuffs. XXIX. Potent protective effects of sesquiterpenes and curcumin from Zedoariae Rhizoma on liver injury induced by D-galactosamine/lipopolysaccharide or tumor necrosis factor-alpha. Biol Pharm Bull 25: 627-31, 2002.
- Ramprasad C and Sirsi M: Studies on Indian medicinal plants: Curcuma longa Linn.- effect of curcumin & the essential oils of C. longa on bile secretion. J Sci Industr Res 15: 262-265, 1956.
- Awasthi S, Srivatava SK, Piper JT, Singhal SS, Chaubey M and Awasthi YC: Curcumin protects against 4-hydroxy-2-trans-nonenal-induced cataract formation in rat lenses. Am J Clin Nutr 64: 761-6, 1996.
- Venkatesan N and Chandrakasan G: Modulation of cyclopho-sphamide-induced early lung injury by curcumin, an anti-inflammatory antioxidant. Mol Cell Biochem 142: 79-87, 1995.
- Venkatesan N, Punithavathi V and Chandrakasan G: Curcumin protects bleomycin-induced lung injury in rats. Life Sci 61: PL51-8, 1997.
- Venkatesan N: Pulmonary protective effects of curcumin against paraquat toxicity. Life Sci 66: PL21-8, 2000.
- Punithavathi D, Venkatesan N and Babu M: Curcumin inhibition of bleomycin- induced pulmonary fibrosis in rats. Br J Pharmacol 131: 169-72, 2000.
- Saleheen D, Ali SA, Ashfaq K, Siddiqui AA, Agha A and Yasinzai MM: Latent activity of curcumin against leishmaniasis in vitro. Biol Pharm Bull 25: 386-9, 2002.
- Gomes Dde C, Alegrio LV, de Lima ME, Leon LL and Araujo CA: Synthetic derivatives of curcumin and their activity against Leishmania amazonensis. Arzneimittelforschung 52: 120-4, 2002.
- Koide T, Nose M, Ogihara Y, Yabu Y and Ohta N: Leishmanicidal effect of curcumin in vitro. Biol Pharm Bull 25: 131-3, 2002.
- Huang HC, Jan TR and Yeh SF: Inhibitory effect of curcumin, an anti-inflammatory agent, on vascular smooth muscle cell proliferation. Eur J Pharmacol 221: 381-4, 1992.
- Chen HW and Huang HC: Effect of curcumin on cell cycle progression and apoptosis in vascular smooth muscle cells. Br J Pharmacol 124: 1029-40, 1998.
- Srimal RC and Dhawan BN: Pharmacology of diferuloylmethane (curcumin), a non- steroidal anti-inflammatory agent. J Pharm Pharmacol 25: 447-52, 1973.
- Conney AH, Lysz T, Ferraro T, Abidi TF, Manchand PS, Laskin JD and Huang MT: Inhibitory effect of curcumin and some related dietary compounds on tumor promotion and arachidonic acid metabolism in mouse skin. Adv Enzyme Regul 31: 385-96, 1991.
- Majeed M, Badmaev V and Murray F: Turmeric and the healing curcuminoids: their amazing antioxidant properties and protective powers. New Canaan: Keats Publishing Inc: 1-46. 1996.
- Surh YJ: Anti-tumor promoting potential of selected spice ingredients with antioxidative and anti-inflammatory activities: a short review. Food Chem Toxicol 40: 1091-7, 2002.
- Bremner P and Heinrich M: Natural products as targeted modulators of the nuclear factor-kappaB pathway. J Pharm Pharmacol 54: 453-72, 2002.
- Gescher AJ, Sharma RA and Steward WP: Cancer chemo-prevention by dietary constituents: a tale of failure and promise. Lancet Oncol 2: 371-9, 2001.
- Chignell CF, Bilski P, Reszka KJ, Motten AG, Sik RH and Dahl TA: Spectral and photochemical properties of curcumin. Photo-chem Photobiol 59: 295-302, 1994.
- Das KC and Das CK: curcumin (diferuloylmethane), a singlet oxygen ((1)0(2)) quencher. Biochem Biophys Res Commun 295: 62-6. 2002.
- Toniolo R, Di Narda F, Susmel S, Martelli M, Martelli L and Bontempelli G: Quenching of superoxide ions by curcumin. A mechanistic study in acetonitrile. Ann Chim 92: 281-8, 2002.
- Dahl TA, Bilski P, Reszka KJ and Chignell CF: Photocytotoxicity of curcumin. Photochem Photobiol 59: 290-4, 1994.
- Tonnesen HH and Karlsen J: Studies on curcumin and curcuminoids. VI. Kinetics of curcumin degradation in aqueous solution. Z Lebensm Unters Forsch 180: 402-4, 1985.
- Tonnesen HH, Karlsen J and van Henegouwen GB: Studies on curcumin and curcuminoids. VIII. Photochemical stability of curcumin. Z Lebensm Unters Forsch 183: 116-22, 1986.
- Tonnesen HH, de Vries H, Karlsen J and Beijersbergen van Henegouwen G: Studies on curcumin and curcuminoids. IX: Investigation of the photobiological activity of curcumin using bacterial indicator systems. J Pharm Sci 76: 371-3, 1987.
- Tonnesen H, Masson M and Loftsson T: Studies of curcumin and curcuminoids. XXVII. Cyclodextrin complexation: solubility, che-mical and photochemical stability. Int J Pharm 244: 127, 2002.
- Wang YJ, Pan MH, Cheng AL, Lin LI, Ho YS, Hsieh CY and Lin JK: Stability of curcumin in buffer solutions and characterization of its degradation products. J Pharm Biomed Anal 15: 1867-76, 1997.
- Chen YC, Kuo TC, Lin-Shiau SY and Lin JK: Induction of HSP70 gene expression by modulation of Ca(+2) ion and cellular p53 protein by curcumin in colorectal carcinoma cells. Mol Carcinog 17: 224-34, 1996.
- Jiang MC, Yang-Yen HF, Yen JJ and Lin JK: Curcumin induces apoptosis in immortalized NIH 3T3 and malignant cancer cell lines. Nutr Cancer 26: 111-20, 1996.
- Kuo ML, Huang TS and Lin JK: Curcumin, an antioxidant and anti-tumor promoter, induces apoptosis in human leukemia cells. Biochim Biophys Acta 1317: 95-100, 1996.
- Mehta K, Pantazis P, McQueen T and Aggarwal BB: Antipro-liferative effect of curcumin (diferuloylmethane) against human breast tumor cell lines. Anticancer Drugs 8: 470-81, 1997.
- Verma SP, Salamone E and Goldin B: Curcumin and genistein, plant natural products, show synergistic inhibitory effects on the growth of human breast cancer MCF-7 cells induced by estrogenic pesticides. Biochem Biophys Res Commun 233: 692-6, 1997.
- Hanif R, Qiao L, Shiff SJ and Rigas B: Curcumin, a natural plant phenolic food additive, inhibits cell proliferation and induces cell cycle changes in colon adenocarcinoma cell lines by prostaglandin-independent pathway. J Lab Clin Med 130: 576-84, 1997.
- Samaha HS, Kelloff GJ, Steele V, Rao CV and Reddy BS: Modulation of apoptosis by sulindac, curcumin, phenylethyl-3-methylcaffeate, and 6-phenylhexyl isothiocyanate: apoptotic index as a biomarker in colon cancer chemoprevention and promotion. Cancer Res 57: 1301-5, 1997.
- Simon A, Allais DP, Duroux JL, Basly JP, Durand-Fontanier S and Delage C: Inhibitory effect of curcuminoids on MCF-7 cell proliferation and structure- activity relationships. Cancer Lett 129: 111-6, 1998.
- Jee SH, Shen SC, Tseng CR, Chiu HC and Kuo ML: Curcumin induces a p53-dependent apoptosis in human basal cell carcinoma cells. J Invest Dermatol 111: 656-61, 1998.
- Nogaki A, Satoh K, Iwasaka K, Takano H, Takahama M, Ida Y and Sakagami H: Radical intensity and cytotoxic activity of curcumin and gallic acid. Anticancer Res 18: 3487-91, 1998.
- Piwocka K, Zablocki K, Wieckowski MR, Skierski J, Feiga I, Szopa J, Drela N, Wojtczak L and Sikora E: A novel apoptosis-like pathway, independent of mitochondria and caspases, induced by curcumin in human lymphoblastoid T (Jurkat) cells. Exp Cell Res 249: 299-307, 1999.
- Khar A, Ali AM, Pardhasaradhi BV, Begum Z and Anjum R: Antitumor activity of curcumin is mediated through the induction of apoptosis in AK-5 tumor cells. FEBS Lett 445: 165-8, 1999.
- Han SS, Chung ST, Robertson DA, Ranjan D and Bondada S: Curcumin causes the growth arrest and apoptosis of B cell lymphoma by downregulation of egr-1, c-myc, bcl-XL, NF-kappa B, and p53. Clin Immunol 93: 152-61, 1999.
- Ramachandran C and You W: Differential sensitivity of human mammary epithelial and breast carcinoma cell lines to curcumin. Breast Cancer Res Treat 54:269-78, 1999.
- Chen H, Zhang ZS, Zhang YL and Zhou DY: Curcumin inhibits cell proliferation by interfering with the cell cycle and inducing apoptosis in colon carcinoma cells. Anticancer Res 19: 3675-80, 1999.
- Bhaumik S, Anjum R, Rangaraj N, Pardhasaradhi BV and Khar A: Curcumin mediated apoptosis in AK-5 tumor cells involves the production of reactive oxygen intermediates. FEBS Lett 456: 311-4, 1999.
- Elattar TM and Virji AS: The inhibitory effect of curcumin, genistein, quercetin and cisplatin on the growth of oral cancer cells in vitro. Anticancer Res 20: 1733-8, 2000.
- Bielak-Zmijewska A, Koronkiewicz M, Skierski J, Piwocka, K, Radziszewska E and Sikora E: Effect of curcumin on the apoptosis of rodent and human nonproliferating and proliferating lymphoid cells. Nutr Cancer 38: 131-8, 2000.
- Cipriani B, Borsellino G, Knowles H, Tramonti D, Cavaliere F, Bernardi G, Battistini L and Brosnan CF: Curcumin inhibits activation of Vgamma9Vdelta2 T cells by phosphoantigens and induces apoptosis involving apoptosis-inducing factor and large scale DNA fragmentation. J Immunol 167: 3454-62, 2001.
- Pan MH, Chang WL, Lin-Shiau SY, Ho CT, Lin JK: Induction of apoptosis by garcinol and curcumin through cytochrome c release and activation of caspases in human leukemia HL-60 cells. J Agric Food Chem 49: 1464-74, 2001.
- Morin D, Barthelemy S, Zini R, Labidalle S and Tillement JP: Curcumin induces the mitochondrial permeability transition pore mediated by membrane protein thiol oxidation. FEBS Lett 495: 131-6, 2001.
- Moragoda L, Jaszewski R and Majumdar AP: Curcumin induced modulation of cell cycle and apoptosis in gastric and colon cancer cells. Anticancer Res 21: 873-8, 2001.
- Mukhopadhyay A, Bueso-Ramos C, Chatterjee D, Pantazis P and Aggarwal BB: Curcumin downregulates cell survival mechanisms in human prostate cancer cell lines. Oncogene 20: 7597-609, 2001.
- Pal S, Choudhuri T, Chattopadhyay S, Bhattacharya A, Datta GK, Das T and Sa G: Mechanisms of curcumin-induced apoptosis of Ehrlich’s ascites carcinoma cells. Biochem Biophys Res Commun 288: 658-65, 2001.
- Kim MS, Kang HJ and Moon A: Inhibition of invasion and induction of apoptosis by curcumin in H-ras-transformed MCF10A human breast epithelial cells. Arch Pharm Res 24: 349-54, 2001.
- Piwocka K, Jaruga E, Skierski J, Gradzka I and Sikora E: Effect of glutathione depletion on caspase-3 independent apoptosis pathway induced by curcumin in Jurkat cells. Free Radic Biol Med 31: 670-8, 2001.
- Bush JA, Cheung KJ, Jr. and Li G: Curcumin induces apoptosis in human melanoma cells through a Fas receptor/caspase-8 pathway independent of p53. Exp Cell Res 271: 305-14, 2001.
- Mori H, Niwa K, Zheng Q, Yamada Y, Sakata K and Yoshimi N: Cell proliferation in cancer prevention; effects of preventive agents on estrogen-related endometrial carcinogenesis model and on an in vitro model in human colorectal cells. Mutat Res 480-481: 201-7,2001.
- Khar A, Ali AM, Pardhasaradhi BV, Varalakshmi CH, Anjum R and Kumari AL: Induction of stress response renders human tumor cell lines resistant to curcumin- mediated apoptosis: role of reactive oxygen intermediates. Cell Stress Chaperones 6: 368-76, 2001.
- Choudhuri T, Pal S, Agwarwal ML, Das T and Sa G: Curcumin induces apoptosis in human breast cancer cells through p53-dependent Bax induction. FEBS Lett 512: 334-40, 2002.
- Anto RJ, Mukhopadhyay A, Denning K and Aggarwal BB: Curcumin (diferuloylmethane) induces apoptosis through activation of caspase-8, BID cleavage and cytochrome c release: its suppression by ectopic expression of Bel-2 and Bcl-xl. Carcinogenesis 23: 143-50, 2002.
- Somasundaram S, Edmund NA, Moore DT, Small GW, Shi YY and Orlowski RZ: Dietary curcumin inhibits chemotherapy-induced apoptosis in models of human breast cancer. Cancer Res 3868-75, 2002.
- Gautam SC, Xu YX, Pindolia KR, Janakiraman N and Chapman RA: Nonselective inhibition of proliferation of transformed and nontransformed cells by the anticancer agent curcumin (diferu-loylmethane). Biochem Pharmacol 55: 1333-7, 1998.
- Ramsewak RS, DeWitt DL and Nair MG: Cytotoxicity, antioxidant and anti- inflammatory activities of curcumins I-III from Curcuma longa. Phytomedicine 7: 303-8, 2000.
- Singh AK, Sidhu GS, Deepa T and Maheshwari RK: Curcumin inhibits the proliferation and cell cycle progression of human umbilical vein endothelial cell. Cancer Lett 107: 109-15, 1996.
- Sup Shim J, Hoon Kim D, Jung HJ, Hee Kim J, Lim D, Lee SK, Kim KW, Ahn JW, Yoo JS, Rho JR, Shin J and Jeong Kwon H: Hydrazinocurcumin, a novel synthetic curcumin derivative, Is a potent inhibitor of endothelial cell proliferation. Bioorg Med Chem 2439-44, 2002.
- Park MJ, Kim EH, Park IC, Lee HC, Woo SH, Lee JY, Hong YJ, Rhee CH, Choi SH, Shim BS, Lee SH and Hong SI: Curcumin inhibits cell cycle progression of immortalized human umbilical vein endothelial (ECV304) cells by up-regulating cyclin-dependent kinase inhibitor, p21 WAF1/CIP1, p27KIP1 and p53. Int J Oncol 379-83, 2002.
- Khafif A, Schantz SP, Chou TC, Edelstein D and Sacks PG: Quantitation of chemopreventive synergism between (-)-epigallo-catechin-3-gallate and curcumin in normal, premalignant and malignant human oral epithelial cells. Carcinogenesis 19: 419-24, 1998.
- Sikora E, Bielak-Zmijewska A, Piwocka K, Skierski J and Radziszewska E: Inhibition of proliferation and apoptosis of human and rat T lymphocytes by curcumin, a curry pigment. Biochem Pharmacol 54: 899-907, 1997.
- Jaruga E, Bielak-Zmijewska A, Sikora E, Skierski J, Radziszewska E, Piwocka K and Bartosz G: Glutathione-independent mechanism of apoptosis inhibition by curcumin in rat thymocytes. Biochem Pharmacol 56: 961-5, 1998.
- Jaruga E, Salvioli S, Dobrucki J, Chrul S, Bandorowicz-Pikula J, Sikora E, Franceschi C, Cossarizza A and Bartosz G: Apoptosis-like, reversible changes in plasma membrane asymmetry and permeability, and transient modifications in mitochondrial membrane potential induced by curcumin in rat thymocytes. FEBS Lett 433: 287-93, 1998.
- Jaruga E, Sokal A, Chrul S and Bartosz G: Apoptosis-independent alterations in membrane dynamics induced by curcumin. Exp Cell Res 245: 303-12, 1998.
- Ozaki K, Kawata Y, Amano S and Hanazawa S: Stimulatory effect of curcumin on osteoclast apoptosis. Biochem Pharmacol 59: 1577-81, 2000.
- Gomez-Lechon MJ, O’Connor E, Castell JV and Jover R: Sensitive markers used to identify compounds that trigger apoptosis in cultured hepatocytes. Toxicol Sci 65: 299-308, 2002.
- Singh S and Aggarwal BB: Activation of transcription factor NF-kappa B is suppressed by curcumin (diferuloylmethane) [corrected]. J Biol Chem 270: 24995- 5000, 1995.
- Xu YX, Pindolia KR, Janakiraman N, Noth CJ, Chapman RA and Gautam SC: Curcumin, a compound with anti-inflammatory and anti-oxidant properties, down-regulates chemokine expression in bone marrow stromal cells. Exp Hematol 2-5: 413-22, 1997.
- Surh YJ, Han SS, Keum YS, Seo HJ and Lee SS: Inhibitory effects of curcumin and capsaicin on phorbol ester-induced activation of eukaryotic transcription factors, NF-κ B and AP-1. Biofactors 12: 107-12, 2000.
- Han SS, Keum YS, Seo HJ and Surh YJ: Curcumin suppresses activation of NF-κ B, and AP-1 induced by phorbol ester in cultured human promyelocytic leukemia cells. J Biochem Mol Biol 35: 337-42, 2002.
- Chuang SE, Yeh PY, Lu YS, Lai GM, Liao CM, Gao M and Cheng AL: Basal levels and patterns of anticancer drug-induced activation of nuclear factor-kappaB (NFkappaB), and its attenuation by tamoxifen, dexamethasone, and curcumin in carcinoma cells. Biochem Pharmacol 63: 1709-16, 2002.
- Chan MM: Inhibition of tumor necrosis factor by curcumin, a phytochemical. Biochem Pharmacol 49: 1551-6, 1995.
- Jang MK, Sohn DH and Ryu JH: A curcuminoid and sesquiterpenes as inhibitors of macrophage TNF-alpha release from Curcuma zedoaria. Planta Med 67: 550-2, 2001.
- Abe Y, Hashimoto S and Horie T: Curcumin inhibition of inflammatory cytokine production by human peripheral blood monocytes and alveolar macrophages. Pharmacol Res 39: 41-7, 1999.
- Literat A, Su F, Norwicki M, Durand M, Ramanathan R, Jones CA, Minoo P and Kwong KY: Regulation of pro-inflammatory cytokine expression by curcumin in hyaline membrane disease (HMD). Life Sci 70: 253-67, 2001.
- Jobin C, Bradham CA, Russo MP, Juma B, Narula AS, Brenner DA and Sartor RB: Curcumin blocks cytokine-mediated NF-kappa B activation and proinflammatory gene expression by inhibiting inhibitory factor I-kappa B kinase activity. J Immunol 163: 3474-83, 1999.
- Hidaka H, Ishiko T, Furuhashi T, Kamohara H, Suzuki S, Miyazaki M, Ikeda O, Mita S, Setoguchi T and Ogawa M: Curcumin inhibits interleukin 8 production and enhances interleukin 8 receptor expression on the cell surface:impact on human pancreatic carcinoma cell growth by autocrine regulation. Cancer 95: 1206-14, 2002.
- Lin LI, Ke YF, Ko YC and Lin JK: Curcumin inhibits SK-Hep- I hepatocellular carcinoma cell invasion in vitro and suppresses matrix metalloproteinase-9 secretion. Oncology 55: 349-53, 1998.
- Mohan R, Sivak J, Ashton P, Russo LA, Pham BQ, Kasahara N, Raizman MB and Fini ME: Curcuminoids inhibit the angiogenic response stimulated by fibroblast growth factor-2, including expression of matrix metalloproteinase gelatinase B. J Biol Chem 2 75: 10405-12, 2000.
- Liacini A, Sylvester J, Li WQ and Zafarullah M: Inhibition of interleukin- 1-stimulated MAP kinases, activating protein-1 (AP-1) and nuclear factor kappa B (NF-kappaB) transcription factors down-regulates matrix metalloproteinase gene expression in articular chondrocytes. Matrix Biol 21: 251-62, 2002.
- Onodera S, Nishihira J, Iwabuchi K, Koyama Y, Yoshida K, Tanaka S and Minami A: Macrophage migration inhibitory factor up-regulates matrix metalloproteinase-9 and -13 in rat osteoblasts. Relevance to intracellular signaling pathways. J Biol Chem 277: 7865-74, 2002.
- Kumar A, Dhawan S, Hardegen NJ and Aggarwal BB: curcumin (Diferuloylmethane) inhibition of tumor necrosis factor (TNF)-mediated adhesion of monocytes to endothelial cells by suppression of cell surface expression of adhesion molecules and of nuclear factor-κ B activation. Biochem Pharmacol 55: 775-83, 1998.
- Gupta B and Ghosh B: Curcuma longa inhibits TNF-alpha induced expression of adhesion molecules on human umbilical vein endothelial cells. Int J Immunopharmacol 21: 745-57, 1999.
- Bierhaus A, Zhang Y, Quehenberger P, Luther T, Haase M, Muller M, Mackman N, Ziegler R and Nawroth PP: The dietary pigment curcumin reduces endothelial tissue factor gene expression by inhibiting binding of AP-1 to the DNA and activation of NF-κ B. Thromb Haemost 77: 772-82, 1997.
- Zhang F, Altorki NK, Mestre JR, Subbaramaiah K and Dannenberg AJ: Curcumin inhibits cyclooxygenase-2 transcription in bile acid- and phorbol ester-treated human gastrointestinal epithelial cells. Carcinogenesis 20: 445-51, 1999.
- Plummer SM, Holloway KA, Manson MM, Munks RJ, Kaptein A, Farrow S and Howells L: Inhibition of cyclo-oxygenase 2 expression in colon cells by the chemopreventive agent curcumin involves inhibition of NF-kappaB activation via the NIK/IKK signalling complex. Oncogene 18: 6013 -20, 1999.
- Goel A, Boland CR and Chauhan DP: Specific inhibition of cyclooxygenase-2 (COX2) expression by dietary curcumin in HT-29 human colon cancer cells. Cancer Lett 172: 111-8, 2001.
- Surh YJ, Chun KS, Cha HH, Han SS, Keum YS, Park KK and Lee Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: down-regulation of COX-2 and iNOS through suppression of NF-kappa B activation. Mutat Res 480-481: 243-68, 2001.
- Chan MM, Huang HI, Fenton MR and Fong D: In vivo inhibition of nitric oxide synthase gene expression by curcumin, a cancer preventive natural product with anti-inflammatory properties. Biochem Pharmacol 55: 1955-62, 1998.
- Brouet I and Ohshima H: Curcumin, an anti-tumour promoter and anti-inflammatory agent, inhibits induction of nitric oxide synthase in activated macrophages. Biochem Biophys Res Commun 206: 533-40, 1995.
- Chan MM, Ho CT and Huang HI: Effects of three dietary phytochemicals from tea, rosemary and turmeric on inflammation-induced nitrite production. Cancer Lett 96: 23-9, 1995.
- Pan MH, Lin-Shiau SY and Lin JK: Comparative studies on the suppression of nitric oxide synthase by curcumin and its hydro-genated metabolites through downregulation of IkappaB kinase and NFkappaB activation in macrophages. Biochem Pharmacol 60: 1665-76, 2000.
- Onoda M and Inano H: Effect of curcumin on the production of nitric oxide by cultured rat mammary gland. Nitric Oxide 4: 505-15, 2000.
- Hour TC, Chen J, Huang CY, Guan JY, Lu SH and Pu YS: Curcumin enhances cytotoxicity of chemotherapeutic agents in prostate cancer cells by inducing p21 (WAF1/CIP1) and C/EBPbeta expressions and suppressing NF-kappaB activation. Prostate 51: 211-8, 2002.
- Huang TS, Lee SC and Lin JK: Suppression of c-Jun/AP-1 activation by an inhibitor of tumor promotion in mouse fibroblast cells. Proc Natl Acad Sci USA 88: 52926, 1991.
- Hanazawa S, Takeshita A, Amano S, Semba T, Nirazuka T, Katoh H and Kitano S: Tumor necrosis factor-alpha induces expression of monocyte chemoattractant JE via fos and jun genes in clonal osteoblastic MC3T3-E1 cells. J Biol Chem 268: 9526-32, 1993.
- Kakar SS and Roy D: Curcumin inhibits TPA induced expression of c-fos, c-jun and c-myc proto-oncogenes messenger RNAs in mouse skin. Cancer Lett 87: 85-9, 1994.
- Lu YP, Chang RL, Lou YR, Huang MT, Newmark HL, Reuhl KR and Conney AH: Effect of curcumin on 12-O-tetradecanoyl-phorbol-13-acetate- and ultraviolet B light-induced expression of c-Jun and c-Fos in J136 cells and in mouse epidermis. Carcinogenesis 15: 2363-70, 1994.
- Xu YX, Pindolia KR, Janakiraman N, Chapman RA and Gautarn SC: Curcumin inhibits IL-1 alpha and TNF-alpha induction of AP-1 and NF-kB DNA-binding activity in bone marrow stromal cells. Hematopathol Mol Hematol 11: 49-62, 1997.
- Chen X, He Q, Liu W, Xu Q, Ye Y, Fu B and Yu L: AP-1 mediated signal transduction in thrombin-induced regulation of PAL-1 expression in human mesangial cells. Chin Med J (Engl) 514-9, 2000.
- Pendurthi UR and Rao LV: Suppression of transcription factor Egr-1 by curcumin. Thromb Res 97: 179-89, 2000.
- Nakamura K, Yasunaga Y, Segawa T, Ko D, Moul JW, Srivastava S and Rhim JS: curcumin down-regulates AR gene expression and activation in prostate cancer cell lines. Int J Oncol 21: 825-30, 2002.
- Korutla L and Kumar R: Inhibitory effect of curcumin on epidermal growth factor receptor kinase activity in A431 cells. Biochim Biophys Acta 1224: 597-600, 1994.
- Korutla L, Cheung JY, Mendelsohn J and Kumar R: Inhibition of ligand-induced activation of epidermal growth factor receptor tyrosine phosphorylation by curcumin. Carcinogenesis 16: 1741-5, 1995.
- Hong RL, Spohn WH and Hung MC: Curcumin inhibits tyrosine kinase activity of p185neu and also depletes p185neu. Clin Cancer Res 5: 1884-91, 1999.
- Dorai T, Gehani N and Katz A: Therapeutic potential of curcumin in human prostate cancer. II. curcumin inhibits tyrosine kinase activity of epidermal growth factor receptor and depletes the protein. Mol Urol 4: 1-6, 2000.
- Liu JY, Lin SJ and Lin JK: Inhibitory effects of curcumin on protein kinase C activity induced by 12-O-tetradecanoyl-phorbol-13-acetate in NIH 3T3 cells. Carcinogenesis 14: 857-61, 1993.
- Reddy S and Aggarwal BB: Curcumin is a non-competitive and selective inhibitor of phosphorylase kinase. FEBS Lett 341: 19-22, 1994.
- Hasmeda M and Polya GM: Inhibition of cyclic AMP-dependent protein kinase by curcumin. Phytochemistry 42: 599-605, 1996.
- Huang MT, Lysz T, Ferraro T, Abidi TF, Laskin JD and Conney AH: Inhibitory effects of curcumin on in vitro lipoxygenase and cyclooxygenase activities in mouse epidermis. Cancer Res 51: 813-9, 1991.
- Skrzypczak-Jankun E, McCabe NP, Selman SH and Jankun J: curcumin inhibits lipoxygenase by binding to its central cavity: theoretical and X-ray evidence. Int J Mol Med 6: 521-6, 2000.
- Susan M and Rao MN: Induction of glutathione S-transferase activity by curcumin in mice. Arzneimittelforschung 42: 962-4, 1992.
- Oetari S, Sudibyo M, Commandeur JN, Samhoedi R and Vermeulen NP: Effects of curcumin on cytochrome P450 and glutathione S-transferase activities in rat liver. Biochem Pharmacol 39-45, 1996.
- Awasthi S, Pandya U, Singhal SS, Lin JT, Thiviyanathan V, Seifert WE, Jr., Awasthi YC and Ansari GA: Curcumin-glutathione interactions and the role of human glutathione S-transferase P1-1. Chem Biol Interact 128: 19-38, 2000.
- Lin JK and Shih CA: Inhibitory effect of curcumin on xanthine dehydrogenase/oxidase induced by phorbol-12-myristate-13-acetate in NIH3 T3 cells. Carcinogenesis 15: 1717-21, 1994.
- Kaul S and Krishnakanth TP: Effect of retinol deficiency and curcumin or turmeric feeding on brain Na(+)-K+ adenosine triphosphatase activity. Mol Cell Biochem 137: 101-7, 1994.
- Thaphyal R and Maru GB: Inhibition of cytochrome P450 isozymes >by curcumins in vitro and in vivo. Food Chem Toxicol 39: 541-7, 2001.
- Ciolino HP, Daschner PJ, Wang TT and Yeh GC: Effect of curcumin on the aryl hydrocarbon receptor and cytochrome P450 1A1 in MCF-7 human breast carcinoma cells. Biochem Pharmacol 197-206, 1998.
- Logan-Smith MJ, Lockyer PJ, East JM and Lee AG: Curcumin, a molecule that inhibits the Ca2+-ATPase of sarcoplasmic reticulum but increases the rate of accumulation of Ca2+. J Biol Chem 276: 46905-11, 2001.
- Logan-Smith MJ, East JM and Lee AG: Evidence for a global inhibitor-induced conformation change on the Ca(2+)-ATPase of sarcoplasmic reticulum, from paired inhibitor studies. Biochemistry 2869-75, 2002.
- Bilmen JG, Khan SZ, Javed MH and Michelangeli F: Inhibition of the SERCA Ca2+ pumps by curcumin. Curcumin putatively stabilizes the interaction between the nucleotide-binding and phosphorylation domains in the absence of ATP. Eur J Biochem 6318-27, 2001.
- Sumbilla C, Lewis D, Hammerschmidt T and Inesi G: The slippage of the Ca2+ pump and its control by anions and curcumin in skeletal and cardiac sarcoplasmic reticulum. J Biol Chem 277: 13900-6, 2002.
- Yamamoto H, Hanada K, Kawasaki K and Nishijima M: Inhibitory effect on curcumin on mammalian phospholipase D activity. FEBS Lett 417: 196-8, 1997.
- Kang BY, Song YJ, Kim KM, Choe YK, Hwang SY and Kim TS: Curcumin inhibits Thl cytokine profile in CD4+ T cells by suppressing interleukin-12 production in macrophages. Br J Pharmacol 128: 380-4, 1999.
- Kang BY, Chung SW, Chung W, Im S, Hwang SY and Kim TS: Inhibition of interleukin-12 production in lipopolysaccharide-activated macrophages by curcumin. Eur J Pharmacol 384: 191-5, 1999.
- Santibanez JF, Quintanilla M and Martinez J: Genistein and curcumin block TGF-beta 1-induced u-PA expression and migratory and invasive phenotype in mouse epidermal kera-tinocytes. Nutr Cancer 37: 49-54, 2000.
- Fenton JI, Wolff MS, Orth MW and Hord NG: Membrane-type matrix metalloproteinases mediate curcumin-induced cell migra-tion in non-tumorigenic colon epithelial cells differing in Apc genotype. Carcinogenesis 23: 1065-70, 2002.
- Motterlini R, Foresti R, Bassi R and Green CJ: Curcumin, an antioxidant and antiinflammatory agent, induces heme oxygenase-1 and protects endothelial cells against oxidative stress. Free Radic Biol Med 28: 1303-12, 2000.
- Scapagnini G, Foresti R, Calabrese V, Giuffrida Stella AM, Green CJ and Motterlini R: Caffeic acid phenethyl ester and curcumin: a novel class of heme oxygenase-1 inducers. Mol Pharmacol 61: 554-61, 2002.
- Romiti N, Tongiani R, Cervelli F and Chieli E: Effects of curcumin on P-glycoprotein in primary cultures of rat hepatocytes. Life Sci 2349-58, 1998.
- Snyder RD and Arnone MR: Putative identification of functional interactions between DNA intercalating agents and topoisomerase II using the V79 in vitro micronucleus assay. Mutat Res 503: 21-35, 2002.
- Kato K, Ito H, Kamei K and Iwamoto I: Stimulation of the stress-induced expression of stress proteins by curcumin in cultured cells and in rat tissues in vivo. Cell Stress Chaperones 3: 152-60, 1998.
- Chen X, Hasuma T, Yano Y, Yoshimata T, Morishima Y, Wang Y and Otani S: Inhibition of farnesyl protein transferase by mono-terpene, curcumin derivatives and gallotannin. Anticancer Res 17: 2555-64, 1997.
- Siebenlist U, Franzoso G and Brown K: Structure, regulation and function of NFkappa B. Annu Rev Cell Biol 10: 405-55, 1994.
- Baeuerle PA and Henkel T: Function and activation of NF-kappa B in the immune system. Annu Rev Immunol 12: 141-79, 1994.
- Grilli M, Chiu JJ and Lenardo MJ: NF-kappa B and Rel: participants in a multiform transcriptional regulatory system. Int Rev Cytol 143: 1-62, 1993.
- Baeuerle PA: The inducible transcription activator NF-kappa B: regulation by distinct protein subunits. Biochim Biophys Acta 1072: 63-80, 1991.
- Brennan P and O’Neill LA: Inhibition of nuclear factor kappaB by direct modification in whole cells–mechanism of action of nordihydroguaiaritic acid, curcumin and thiol modifiers. Biochem Pharmacol 55: 965-73, 1998.
- Ishii H, Takada K, Higuchi T and Sugiyama J: Verotoxin-1 induces tissue factor expression in human umbilical vein endothelial cells through activation of NFkappaB/Rel and AP-1. Thromb Haemost 84: 712-21, 2000.
- Anto RJ, Maliekal TT and Karunagaran D: L-929 cells harboring ectopically expressed ReIA resist curcumin-induced apoptosis. J Biol Chem 275: 15601-4, 2000.
- Bharti AC, Donato N, Singh S and Aggarwal BB: Curcumin (diferuloylmethane) downregulates the constitutive activation of nuclear factor-kappaB and I kappa B alpha kinase in human multiple myeloma cells leading to suppression of proliferation and induction of apoptosis. Blood (in press).
- Aggarwal BB: Apoptosis and transcription factor NF-kB: a tale of association and dissociation. Biochem Pharmacol 60: 1033-1039, 2000.
- Shishodia S and Aggrwal BB: Nuclear factor-kappa B activation: a question of life and death. J Biochem Mol Biol 35: 28-40, 2002.
- Karunagaran D and Aggarwal BB: Transcription factors as targets for the drug development. In: Keri G, Toth I, eds. Molecular Pathomechanisms and New Trends in Drug Development. New York: Harwood Academic Publisher, 2002: 76-91.
- Dhar A, Young MR, Colburn NH: The role of AP-1, NF-kappaB and ROS/NOS in skin carcinogenesis: the J136 model is predictive.
- Mol Cell Biochem 234235: 185-93, 2002. Shaulian E and Karin M: AP-1 as a regulator of cell life and death.
- Nat Cell Biol 4: E 131-6, 2002. Hsu TC, Young MR, Cmarik J and Colburn NH: Activator protein 1 (AP-1)- and nuclear factor kappaB (NF-kappaB)-dependent transcriptional events in carcinogenesis. Free Radic Biol Med 28: 1338-48, 2000.
- Dong Z, Lavrovsky V and Colburn NH: Transformation reversion induced in J136 RT101 cells by AP-1 inhibitors. Carcinogenesis 16: 749-56, 1995.
- Chen YR, Zhou G and Tan TH: c-Jun N-terminal kinase mediates apoptotic signaling induced by N-(4-hydroxyphenyl)retinamide. Mol Pharmacol 56: 1271-9, 1999.
- Hahm ER, Cheon G, Lee J, Kim B, Park C and Yang CH: New and known symmetrical curcumin derivatives inhibit the formation of Fos-Jun-DNA complex. Cancer Lett 184: 89-96, 2002.
- Ammon HP, Safayhi H, Mack T and Sabieraj J: Mechanism of antiinflammatory actions of curcumin and boswellic acids. J Ethnopharmacol 38: 113-9, 1993.
- Sherr CJ: D-type cyclins. Trends Biochem Sci 20: 187-90, 1995.
- Baldin V, Lukas J, Marcote MJ, Pagano M and Draetta G: Cyclin D1 is a nuclear protein required for cell cycle progression in G1. Genes Dev 7: 812-21, 1993.
- Bartkova J, Lukas J, Strauss M and Bartek J: The PRAD-1/cyclin D1 oncogene product accumulates aberrantly in a subset of colorectal carcinomas. Int J Cancer 58: 568-73, 1994.
- Bartkova J, Lukas J, Muller H, Lutzhoft D, Strauss M and Bartek J: Cyclin D1 protein expression and function in human breast cancer. Int J Cancer 57: 353-61, 1994.
- Gillett C, Fantl V, Smith R, Fisher C, Bartek J, Dickson C, Barnes D and Peters G: Amplification and overexpression of cyclin D1 in breast cancer detected by immunohistochemical staining. Cancer Res 54: 1812-7, 1994.
- Nishida N, Fukuda Y, Komeda T, Kita R, Sando T, Furukawa M, Amenomori M, Shibagaki I, Nakao K, Ikenaga M et al.: Amplification and overexpression of the cyclin D1 gene in aggressive human hepatocellular carcinoma. Cancer Res 54: 3107-10, 1994.
- Dickson C, Fantl V, Gillett C, Brookes S, Bartek J, Smith R, Fisher C, Barnes D and Peters G: Amplification of chromosome band 11q13 and a role for cyclin D1 in human breast cancer. Cancer Lett 43-50, 1995.
- Bartkova J, Lukas J, Muller H, Strauss M, Gusterson B and Bartek J: Abnormal patterns of D-type cyclin expression and GI regulation in human head and neck cancer. Cancer Res 55: 949-56, 1995.
- Adelaide J, Monges G, Derderian C, Seitz JF and Birnbaum D: Oesophageal cancer and amplification of the human cyclin D gene CCND1/PRAD1. Br J Cancer 71: 64-8, 1995,
- Arber N, Hibshoosh H, Moss SF, Sutter T, Zhang Y, Begg M, Wang S, Weinstein IB and Holt PR: Increased expression of cyclin D1 is an early event in multistage colorectal carcinogenesis. Gastro-enterology 110: 669-74, 1996.
- Caputi M, Groeger AM, Esposito V, Dean C, De Luca A, Pacilio C, Muller MR, Giordano GG, Baldi F, Wolner E and Giordano A: Prognostic role of cyclin D1 in lung cancer. Relationship to proliferating cell nuclear antigen. Am J Respir Cell Mol Biol 20: 746-50, 1999.
- Gumbiner LM, Gumerlock PH, Mack PC, Chi SG, deVere White RW, Mohler JL, Pretlow TG and Tricoli JV: Overexpression of cyclin D1 is rare in human prostate carcinoma. Prostate 38: 40-5, 1999.
- Drobnjak M, Osman I, Scher HI, Fazzari M and Cordon-Cardo C: Overexpression of cyclin D1 is associated with metastatic prostate cancer to bone. Clin Cancer Res 6: 1891-5, 2000.
- Harari D and Yarden Y: Molecular mechanisms underlying ErbB2/HER2 action in breast cancer. Oncogene 19: 6102-14, 2000.
- Wang TC, Cardiff RD, Zukerberg L, Lees E, Arnold A and Schmidt EV: Mammary hyperplasia and carcinoma in MMTV-cyclin D1 transgenic mice. Nature 369: 66971, 1994.
- Deane NG, Parker MA, Aramandla R, Diehl L, Lee WJ, Washington MK, Nanney LB, Shyr Y and Beauchamp RD: Hepatocellular carcinoma results from chronic cyclin D1 overexpression in transgenic mice. Cancer Res 61: 5389-95, 2001.
- Fantl V, Stamp G, Andrews A, Rosewell I and Dickson C: Mice lacking cyclin D1 are small and show defects in eye and mammary gland development. Genes Dev 9: 2364-72, 1995.
- Sicinski P and Weinberg RA: A specific role for cyclin D1 in mammary gland development. J Mammary Gland Biol Neoplasia 2: 335-42, 1997.
- Fantl V, Creer A, Dillon C, Bresnick J, Jackson D, Edwards P, Rosewell I and Dickson C: Fibroblast growth factor signalling and cyclin D1 function are necessary for normal mammary gland development during pregnancy. A transgenic mouse approach. Adv Exp Med Biol 480: 1-7, 2000.
- Yu Q, Geng Y and Sicinski P: Specific protection against breast cancers by cyclin D1 ablation. Nature 411: 1017-21, 2001.
- Albanese C, Johnson J, Watanabe G, Eklund N, Vu D, Arnold A and Pestell RG: Transforming p21 ras mutants and c-Ets-2 activate the cyclin D1 promoter through distinguishable regions. J Biol Chem 270: 23589-97, 1995.
- Joyce D, Bouzahzah B, Fu M, Albanese C, D’Amico M, Steer J, Klein JU, Lee RJ, Segall JE, Westwick JK, Der CJ and Pestell RG: Integration of Rac-dependent regulation of cyclin D1 transcription through a nuclear factor-kappaB-dependent pathway. J Biol Chem 25245-9, 1999.
- Lee RJ, Albanese C, Fu M, D’Amico M, Lin B, Watanabe G, Haines GK, 3rd, Siegel PM, Hung MC, Yarden Y, Horowitz JM, Muller WJ and Pestell RG: Cyclin D1 is required for transformation by activated Neu and is induced through an E2F-dependent signaling pathway. Mol Cell Biol 20: 672-83, 2000.
- Sauter ER, Nesbit M, Litwin S, Klein-Szanto AJ, Cheffetz S and Herlyn M: Antisense cyclin D1 induces apoptosis and tumor shrinkage in human squamous carcinomas. Cancer Res 59: 4876-81, 1999.
- Mukhopadhyay A, Banerjee S, Stafford LJ, Xia C, X., Liu M and Aggarwal BB: Curcumin-induced suppression of cell proliferation correlates with donregulation of cyclin D1 expression and CDK4-mediated retinoblastoma protein phosphorylation. Oncogene (in press).
- Aggarwal BB and Natarajan K: Tumor necrosis factor: developments during the last dacade. Eur Cytokine Netw 7: 93-124, 1996.
- Pober JS and Cotran RS: Cytokines and endothelial cell biology.Physiol Rev 70: 427- 51, 1990.
- Grau GE and Lou J: TNF in vascular pathology: the importance of plateletendothelium interactions. Res Immunol 144: 355-63, 1993.
- Collins T, Read MA, Neish AS, Whitley MZ, Thanos D and Maniatis T: Transcriptional regulation of endothelial cell adhesion molecules: NF-kappa B and cytokine-inducible enhancers. Faseb J 9: 899-909, 1995.
- Thaloor D, Singh AK, Sidhu GS, Prasad PV, Kleinman HK and Maheshwari RK: Inhibition of angiogenic differentiation of human umbilical vein endothelial cells by curcumin. Cell Growth Differ 9: 305-12, 1998.
- Shin EY, Kim SY and Kim EG: c-Jun N-terminal kinase is involved in motility of endothelial cell. Exp Mol Med 33: 276-83, 2001.
- Arbiser JL, Klauber N, Rohan R, van Leeuwen R, Huang MT, Fisher C, Flynn E and Byers HR: Curcumin is an in vivo inhibitor of angiogenesis. Mol Med 4: 376-83, 1998.
- Anuchapreeda S, Leechanachai P, Smith MM, Ambudkar SV and Limtrakul PN: Modulation of P-glycoprotein expression and function by curcumin in multidrugresistant human KB cells. Biochem Pharmacol 64: 573-82, 2002.
- lersel ML, Ploemen JP, Struik I, van Amersfoort C, Keyzer AE, Schefferlie JG and van Bladeren PJ: Inhibition of glutathione S-transferase activity in human melanoma cells by alpha, beta-unsaturated carbonyl derivatives. Effects of acrolein, cinna-maldehyde, citral, crotonaldehyde, curcumin, ethacrynic acid, and trans-2-hexenal. Chem Biol Interact 102: 117-32, 1996.
- Rao TS, Basu N and Siddiqui HH: Anti-inflammatory activity of curcumin analogues. Indian J Med Res 75: 574-8, 1982.
- Mukhopadhyay A, Basu N, Ghatak N and Gujral PK: Anti-inflammatory and irritant activities of curcumin analogues in rats. Agents Actions 12: 508-15, 1982.
- Srivastava R and Srimal RC: Modification of certain inflammation-induced biochemical changes by curcumin. Indian J Med Res 81: 215-23, 1985.
- Reddy AC and Lokesh BR: Studies on anti-inflammatory activity of spice principles and dietary n-3 polyunsaturated fatty acids on carrageenan-induced inflammation in rats. Ann Nutr Metab 38: 349-58, 1994.
- Joe B and Lokesh BR: Effect of curcumin and capsaicin on arachidonic acid metabolism and lysosomal enzyme secretion by rat peritoneal macrophages. Lipids 32: 1173-80, 1997.
- Nagabhushan M, Amonkar AJ and Bhide SV: In vitro antimu-tagenicity of curcumin against environmental mutagens. Food Chem Toxicol 25: 545-7, 1987.
- Shalini VK and Srinivas L: Lipid peroxide induced DNA damage: protection by turmeric (Curcuma longa). Mol Cell Biochem 77: 3-10, 1987.
- Shalini VK and Srinivas L: Fuel smoke condensate induced DNA damage in human lymphocytes and protection by turmeric (Curcuma longa). Mol Cell Biochern 95: 21-30, 1990.
- Donatus IA, Sardjoko and Vermeulen NP: Cytotoxic and cytoprotective activities of curcumin. Effects on paracetamol-induced cytotoxicity, lipid peroxidation and glutathione depletion in rat hepatocytes. Biochem Pharmacol 39: 1869-75, 1990.
- Srinivas L and Shalini VK: DNA damage by smoke: protection by turmeric and other inhibitors of ROS. Free Radic Biol Med 11: 277-83, 1991.
- Azuine MA, Kayal JJ and Bhide SV: Protective role of aqueous turmeric extract against mutagenicity of direct-acting carcinogens as well as benzo [alpha] pyrene-induced genotoxicity and carcinogenicity. J Cancer Res Clin Oncol 118: 447-52, 1992.
- Abraham SK, Sarma L and Kesavan PC: Protective effects of chlorogenic acid, curcumin and beta-carotene against gamma-radiation-induced in vivo chromosomal damage. Mutat Res 303: 109-12, 1993.
- Soudamini KK, Unnikrishnan MC, Sukumaran K and Kuttan R: Mutagenicity and anti-mutagenicity of selected spices. Indian J Physiol Pharmacol 39: 347-53, 1995.
- Oda Y: Inhibitory effect of curcumin on SOS functions induced by UV irradiation. Mutat Res 348: 67-73, 1995.
- Deshpande SS and Maru GB: Effects of curcumin on the formation of benzo[a]pyrene derived DNA adducts in vitro. Cancer Lett 96: 71-80, 1995.
- Anto RJ, George J, Babu KV, Rajasekharan KN and Kuttan R: Antimutagenic and anticarcinogenic activity of natural and synthetic curcuminoids. Mutat Res 370: 127-31, 1996.
- Soni KB, Lahiri M, Chackradeo P, Bhide SV and Kuttan R: Protective effect of food additives on aflatoxin-induced muta – genicity and hepatocarcinogenicity. Cancer Lett 115: 129 – 33, 1997.
- Ahsan H and Hadi SM: Strand scission in DNA induced by curcumin in the presence of Cu(II). Cancer Lett 124: 23-30, 1998.
- Antunes LM, Araujo MC, Dias FL and Takahashi CS: Modulatory effects of curcumin on the chromosomal damage induced by doxorubicin in Chinese hamster ovary cells. Teratog Carcinog Mutagen 19: 1-8, 1999.
- Kelly MR, Xu J, Alexander KE and Loo G: Disparate effects of similar phenolic phytochemicals as inhibitors of oxidative damage to cellular DNA. Mutat Res 485: 309-18, 2001.
- Araujo MC, Antunes LM and Takahashi CS: Protective effect of thiourea, a hydroxyl-radical scavenger, on curcumin-induced chromosomal aberrations in an in vitro mammalian cell system. Teratog Carcinog Mutagen 21: 175-80, 2001.
- Holy JM: Curcumin disrupts mitotic spindle structure and induces micronucleation in MCF-7 breast cancer cells. Mutat Res 518: 71-84, 2002.
- Shukla Y, Arora A and Taneja P: Antimutagenic potential of curcumin on chromosomal aberrations in Wistar rats. Mutat Res 515: 197-202, 2002.
- Grandjean-Laquerriere A, Gangloff SC, Le Naour R, Trentesaux C, Hornebeck W and Guenounou M: Relative contribution of NF-kappaB and AP-1 in the modulation by curcumin and pyrrolidine dithiocarbate of the UVB-induced cytokine expression by kerati-nocytes. Cytokine 18: 168-77, 2002.
- Thresiamma KC, George J and Kuttan R: Protective effect of curcumin, ellagic acid and bixin on radiation induced toxicity. Indian J Exp Biol 34: 845-7, 1996.
- Thresiamma KC, George J and Kuttan R: Protective effect of curcumin, ellagic acid and bixin on radiation induced genotoxicity. J Exp Clin Cancer Res 17: 431-4, 1998.
- Araujo MC, Dias FL and Takahashi CS: Potentiation by turmeric and curcumin of gamma-radiation-induced chromosome aberrations in Chinese hamster ovary cells. Teratog Carcinog Mutagen 19:9-18, 1999.
- Kapoor S and Priyadarsini KI: Protection of radiation-induced protein damage by curcumin. Biophys Chem 92: 119-26, 2001.
- Varadkar P, Dubey P, Krishna M and Verma N: Modulation of radiation-induced protein kinase C activity by phenolics. J Radiol Prot 21: 361-70, 2001.
- Oguro T and Yoshida T: Effect of ultraviolet A on ornithine decarboxylase and metallothionein gene expression in mouse skin. Photodermatol Photoimmunol Photomed 17: 71-8, 2001.
- Ramachandran C, Fonseca HB, Jhabvala P, Escalon EA and Melnick SJ: Curcumin inhibits telomerase activity through human telomerase reverse transcritpase in MCF-7 breast cancer cell line. Cancer Lett 184: 1-6, 2002.
- Holder GM, Plummer JL and Ryan AJ: The metabolism and excretion of curcumin (1,7-bis-(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) in the rat. Xenobiotica 8: 761-8, 1978.
- Wahlstrom B and Blennow G: A study on the fate of curcumin in the rat. Acta Pharmacol Toxicol (Copenh) 43: 86-92, 1978.
- Ravindranath V and Chandrasekhara N: Absorption and tissue distribution of curcumin in rats. Toxicology 16: 259-65, 1980.
- Ravindranath V and Chandrasekhara N: In vitro studies on the intestinal absorption of curcumin in rats. Toxicology 20: 251-7, 1981.
- Ravindranath V and Chandrasekhara N: Metabolism of curcumin– studies with [3H]curcumin. Toxicology 22: 337-44, 1981.
- Shoba G, Joy D, Joseph T, Majeed M, Rajendran R and Srinivas PS: Influence of piperine on the pharmacokinetics of curcumin in animals and human volunteers. Planta Med 64: 353-6, 1998.
- Pan MH, Huang TM and Lin JK: Biotransformation of curcumin through reduction and glucuronidation in mice. Drug Metab Dispos 27: 486-94, 1999.
- Mohandas KM and Desai DC: Epidemiology of digestive tract cancers in India. V. Large and small bowel. Indian J Gastroenterol 118-21, 1999.
- Lal B, Kapoor AK, Asthana OP, Agrawal PK, Prasad R, Kumar P and Srimal RC: Efficacy of curcumin in the management of chronic anterior uveitis. Phytother Res 13: 318-22, 1999.
- Thaloor D, Miller KJ, Gephart J, Mitchell PO and Pavlath GK: Systemic administration of the NF-kappaB inhibitor curcumin stimulates muscle regeneration after traumatic injury. Am J Physiol C320-9, 1999.
- Lin JK, Pan MH and Lin-Shiau SY: Recent studies on the biofunctions and biotransformations of curcumin. Biofactors 13: 153-8, 2000.
- Lal B, Kapoor AK, Agrawal PK, Asthana OP and Srimal RC: Role of curcumin in idiopathic inflammatory orbital pseudotumours. Phytother Res 14: 443-7, 2000.
- Plummer SM, Hill KA, Festing MF, Steward WP, Gescher AJ and Sharma RA: Clinical development of leukocyte cyclooxygenase 2 activity as a systemic biomarker for cancer chemopreventive agents. Cancer Epidemiol Biomarkers Prev 10: 1295-9, 2001.
- Cheng AL, Hsu CH, Lin JK, Hsu MM, Ho YF, Shen TS, Ko JY, Lin JT, Lin BR, Ming-Shiang W, Yu HS, Jee SH, Chen GS, Chen TM, Chen CA, Lai MK, Pu YS, Pan MH, Wang YJ, Tsai CC and Hsieh CY: Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Res 21: 2895-900, 2001.
- Sharma RA, McLelland HR, Hill KA, Ireson CR, Euden SA, Manson MM, Pirmohamed M, Marnett LJ, Gescher AJ and Steward WP: Pharmacodynamic and pharmacokinetic study of oral Curcuma extract in patients with colorectal cancer. Clin Cancer Res 1894-900, 2001.
- Ireson C, Orr S, Jones DJ, Verschoyle R, Lim CK, Luo JL, Howells L, Plummer S, Jukes R, Williams M, Steward WP and Gescher A: Characterization of metabolites of the chemopreventive agent curcumin in human and rat hepatocytes and in the rat in vivo, and evaluation of their ability to inhibit phorbol ester-induced prostaglandin E2 production. Cancer Res 61: 1058-64, 2001.
- Ireson CR, Jones DJ, Orr S, Coughtrie MW, Boocock DJ, Williams ML, Fanner PB, Steward WP and Gescher AJ: Metabolism of the cancer chemopreventive agent curcumin in human and rat intestine. Cancer Epidemiol Biomarkers Prev 11: 10511,2002.
- Hsu CH, Chuang SE, Hergenhahn M, Kuo ML, Lin JK, Hsieh CY and Cheng AL: Pre-clinical and early-phase clinical studies of curcumin as chemopreventive agent for endemic cancers in Taiwan. Gan To Kagaku Ryoho 29 Suppl 1: 194-200, 2002.
- Sharma RA, Ireson CR, Verschoyle RD, Hill KA, Williams ML, Leuratti C, Manson MM, Marnett LJ, Steward WP and Gescher A: Effects of dietary curcumin on glutathione S-transferase and malondialdehyde-DNA adducts in rat liver and colon mucosa: relationship with drug levels. Clin Cancer Res 7: 1452-8, 2001.
- Perkins S, Verschoyle RD, Hill K, Parveen π, Threadgill MD, Sharma RA, Williams ML, Steward WP and Gescher AJ: Chemopreventive efficacy and pharmacokinetics of curcumin in the min/+ mouse, a model of familial adenomatous polyposis. Cancer Epiderniol Biomarkers Prev 11: 535-40, 2002.
- Kuttan R, Bhanumathy P, Nirmala K and George MC: Potential anticancer activity of turmeric (Curcuma longa). Cancer Lett 29: 197-202, 1985.
- Kuttan R, Sudheeran PC and Josph CD: Turmeric and curcumin as topical agents in cancer therapy. Tumori 73: 29-31, 1987.
- Huang MT, Smart RC, Wong CQ and Conney AH: Inhibitory effect of curcumin, chlorogenic acid, caffeic acid, and ferulic acid on tumor promotion in mouse skin by 12-O-tetradecanoylphorbol-13-acetate. Cancer Res 48: 5941-6, 1988.
- Soudamini KK and Kuttan R: Inhibition of chemical carcinogenesisby curcumin. J Ethnopharmacol 27: 227-33, 1989.
- Soni KB, Rajan A and Kuttan R: Reversal of aflatoxin induced liver damage by turmeric and curcumin. Cancer Lett 66: 115-21, 1992.
- Huang MT, Wang ZY, Georgiadis CA, Laskin JD and Conney AH: Inhibitory effects of curcumin on tumor initiation by benzo [a]pyrene and 7,12dimethylbenz[a]anthracene. Carcinogenesis 13: 2183-6, 1992.
- Huang MT, Lou YR, Ma W, Newmark HL, Reuhl KR and Conney AH: Inhibitory effects of dietary curcumin on forestomach, duodenal, and colon carcinogenesis in mice. Cancer Res 54: 5841-7, 1994.
- Nagabhushan M and Bhide SV: Curcumin as an inhibitor of cancer. J Am Coll Nutr 11: 192-8, 1992.
- Azuine MA and Bhide SV: Chemopreventive effect of turmeric against stomach and skin tumors induced by chemical carcinogens in Swiss mice. Nutr Cancer 17: 77-83, 1992.
- Azuine MA and Bhide SV: Protective single/combined treatment with betel leaf and turmeric against methyl (acetoxymethyl) nitrosamine-induced hamster oral carcinogenesis. Int J Cancer 51: 412-5, 1992.
- Rao CV, Simi B and Reddy BS: Inhibition by dietary curcumin of azoxymethane-induced ornithine decarboxylase, tyrosine protein kinase, arachidonic acid metabolism and aberrant crypt foci formation in the rat colon. Carcinogenesis 14: 2219-25, 1993.
- Lu YP, Chang RL, Huang MT and Conney AH: Inhibitory effect of curcumin on 120-tetradecanoylphorbol-13-acetate-induced increase in ornithine decarboxylase mRNA in mouse epidermis. Carcinogenesis 14: 293-7, 1993.
- Tanaka T, Makita H, Ohnishi M, Hirose Y, Wang A, Mori H, Satoh K, Hara A and Ogawa H: Chemoprevention of 4-nitro-quinoline 1-oxide-induced oral carcinogenesis by dietary curcumin and hesperidin: comparison with the protective effect of beta-carotene. Cancer Res 54: 4653-9, 1994.
- Azuine MA and Bhide SV: Adjuvant chemoprevention of expe-rimental cancer: catechin and dietary turmeric in forestomach and oral cancer models. J Ethnopharmacol 44: 211-7, 1994.
- Ghaisas SID and Bhide SV: In vitro studies on chemoprotective effect of Pumark against benzo(a)pyrene-induced chromosomal damage in human lymphocytes. Cell Biol Int 18: 21-7, 1994.
- Rao CV, Rivenson A, Simi B and Reddy BS: Chemoprevention of colon carcinogenesis by dietary curcumin, a naturally occurring plant phenolic compound. Cancer Res 55: 259-66, 1995.
- Rao CV, Rivenson A, Simi B and Reddy BS: Chemoprevention of colon cancer by dietary curcumin. Ann N Y Acad Sci 768: 201-4, 1995.
- Huang MT, Ma W, Lu YP, Chang RL, Fisher C, Manchand PS, Newmark HL and Conney AH: Effects of curcumin, demethoxycurcumin, bisdemethoxycurcumin and tetrahydrocurcumin on 12-O-tetradecanoylphorbol-13-acetate-induced tumor promotion. Carcinogenesis 16: 2493-7, 1995.
- Pereira MA, Grubbs CJ, Barnes LH, Li H, Olson GR, Eto I, Juliana M, Whitaker LM, Kelloff GJ, Steele VE and Lubet RA: Effects of the phytochemicals, curcumin and quercetin, upon azoxy-methane-induced colon cancer and 7,12dimethylbenz[a] anthra-cene-induced mammary cancer in rats. Carcinogenesis 17: 1305-11, 1996.
- Singletary K, MacDonald C, Wallig M and Fisher C: Inhibition of 7,12dimethylbenz[a]anthracene (DMBA)-induced mammary tumo-rigenesis and DMBA-DNA adduct formation by curcumin. Cancer Lett 103: 137-41, 1996.
- Ishizaki C, Oguro T, Yoshida T, Wen CQ, Sueki H and Iijima M: Enhancing effect of ultraviolet A on ornithine decarboxylase induction and dermatitis evoked by 12-otetradecanoylphorbol-13-acetate and its inhibition by curcumin in mouse skin. Dermatology 193: 311-7, 1996.
- Huang MT, Ma W, Yen P, Xie JG, Han J, Frenkel K, Grunberger D and Conney AH: Inhibitory effects of topical application of low doses of curcumin on 12-O-tetradecanoylphorbol-13-acetate-induced tumor promotion and oxidized DNA bases in mouse epidermis. Carcinogenesis 18: 83-8, 1997.
- Limtrakul P, Lipigorngoson S, Namwong O, Apisariyakul A and Dunn FW: Inhibitory effect of dietary curcumin on skin carcinogenesis in mice. Cancer Lett 116: 197-203, 1997.
- Takaba K, Hirose M, Yoshida Y, Kimura J, Ito N and Shirai T: Effects of ntritriacontane-16,18-dione, curcumin, chlorphyllin, dihydroguaiaretic acid, tannic acid and phytic acid on the initiation stage in a rat multi-organ carcinogenesis model. Cancer Lett 113: 39-46, 1997.
- Deshpande SS, Ingle AD and Maru GB: Chemopreventive efficacy of curcumin-free aqueous turmeric extract in 7,12-dimethylbenz [a]anthracene-induced rat mammary tumorigenesis. Cancer Lett 123: 35-40, 1998.
- Singh SV, Hu X, Srivastava SK, Singh M, Xia H, Orchard JL and Zaren HA: Mechanism of inhibition of benzo[a]pyrene-induced forestomach cancer in mice by dietary curcumin. Carcinogenesis 19: 1357-60, 1998.
- Krishnaswamy K, Goud VK, Sesikeran B, Mukundan MA and Krishna TP: Retardation of experimental tumorigenesis and reduction in DNA adducts by turmeric and curcumin. Nutr Cancer 30: 163-6, 1998.
- Piper JT, Singhal SS, Salameh MS, Torman RT, Awasthi YC and Awasthi S: Mechanisms of anticarcinogenic properties of curcumin: the effect of curcumin on glutathione linked detoxification enzymes in rat liver. Int J Biochem Cell Biol 30: 445-56, 1998.
- Kim JM, Araki S, Kim DJ, Park CB, Takasuka N, Baba-Toriyama H, Ota T, Nir Z, Khachik F, Shimidzu N, Tanaka Y, Osawa T, Uraji T, Murakoshi M, Nishino H and Tsuda H: Chemopreventive effects of carotenoids and curcumins on mouse colon carcino-genesis after 1,2-dimethylhydrazine initiation. Carcinogenesis 19: 81-5, 1998.
- Huang MT, Lou YR, Xie JG, Ma W, Lu YP, Yen P, Zhu BT, Newmark H and Ho CT: Effect of dietary curcumin and dibenzoylmethane on formation of 7,12 dimethylbenz[a]anthracene-induced mammary tumors and lymphomas/leukemias in Sencar mice. Carcinogenesis 19: 1697-700, 1998.
- Kawamori T, Lubet R, Steele VE, Kelloff GJ, Kaskey RB, Rao CV and Reddy BS: Chemopreventive effect of curcumin, a naturally occurring anti-inflammatory agent, during the promotion/ progression stages of colon cancer. Cancer Res 59: 597-601, 1999.
- Inano H, Onoda M, Inafuku N, Kubota M, Kamada Y, Osawa T, Kobayashi H and Wakabayashi K: Chemoprevention by curcumin during the promotion stage of tumorigenesis of mammary gland in rats irradiated with gamma-rays. Carcinogenesis 20: 1011-8, 1999.
- Lee SK, Pezzuto JM: Evaluation of the potential of cancer chemo-preventive activity mediated by inhibition of 12-O-tetradecanoyl phorbol 13-acetate-induced ornithine decarboxylase activity. Arch Pharm Res 22: 559-64, 1999.
- Navis I, Sriganth P and Premalatha B: Dietary curcumin with cisplatin administration modulates tumour marker indices in experimental fibrosarcoma. Pharmacol Res 39: 175-9, 1999.
- Chun KS, Sohn Y, Kim HS, Kim OH, Park KK, Lee JM, Moon A, Lee SS and Surh YJ: Anti-tumor promoting potential of naturally occurring diarylheptanoids structurally related to curcumin. Mutat Res 428: 49-57, 1999.
- Hecht SS, Kenney PM, Wang M, Trushin N, Agarwal S, Rao AV and Upadhyaya P: Evaluation of butylated hydroxyanisole, myo-inositol, curcumin, esculetin, resveratrol and lycopene as inhibitors of benzo[a]pyrene plus 4(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced lung turnorigenesis in A/J mice. Cancer Lett 137: 123-30, 1999.
- Churchill M, Chadbum A, Bilinski RT and Bertagnolli MM: Inhibition of intestinal tumors by curcumin is associated with changes in the intestinal immune cell profile. J Surg Res 89: 169-75, 2000.
- Chuang SE, Kuo ML, Hsu CH, Chen CR, Lin JK, Lai GM, Hsieh CY and Cheng AL: Curcumin-containing diet inhibits diethylnitro-samine-induced murine hepatocarcinogenesis. Carcinogenesis 21: 331-5, 2000.
- Chuang SE, Cheng AL, Lin JK and Kuo ML: Inhibition by curcumin of diethylnitrosamine-induced hepatic hyperplasia, inflammation, cellular gene products and cell-cycle-related proteins in rats. Food Chem Toxicol 38: 991-5, 2000.
- Mariadason JM, Comer GA and Augenlicht LH: Genetic repro-gramming in pathways of colonic cell maturation induced by short chain fatty acids: comparison with trichostatin A, sulindac, and curcumin and implications for chemoprevention of colon cancer. Cancer Res 60: 4561-72, 2000.
- Ushida J, Sugie S, Kawabata K, Pham QV, Tanaka T, Fujii K, Takeuchi H, Ito Y and Mori H: Chemopreventive effect of curcumin on N-nitrosomethylbenzylamine-induced esophageal carcinogenesis in rats. Jpn J Cancer Res 91: 893-8, 2000.
- Inano H, Onoda M, Inafuku N, Kubota M, Kamada Y, Osawa T, Kobayashi H and Wakabayashi K: Potent preventive action of curcumin on radiation-induced initiation of mammary tumorige-nesis in rats. Carcinogenesis 21: 1835-41, 2000.
- Lee SE, Campbell BC, Molyneux RJ, Hasegawa S and Lee HS: Inhibitory effects of naturally occurring compounds on aflatoxin B(1) biotransformation. J Agric Food Chem 49: 5171-7, 2001.
- Thaphyal R, Deshpande SS and Maru GB: Effects of turmeric on the activities of benzo(a)pyrene-induced cytochrome P-450 iso-zymes. J Environ Pathol Toxicol Oncol 20: 59-63, 2001.
- Ikezaki S, Nishikawa A, Furukawa F, Kudo K, Nakamura H, Tamura K and Mori H: Chemopreventive effects of curcumin on glandular stomach carcinogenesis induced by N-methyl-N’-nitro-N-nitrosoguanidine and sodium chloride in rats. Anticancer Res 21: 3407-11, 2001.
- Lin CC, Ho CT and Huang MT: Mechanistic studies on the inhibitory action of dietary dibenzoylmethane, a beta-diketone analogue of curcumin, on 7,12dimethylbenz[a]anthracene-induced mammary tumorigenesis. Proc Natl Sci Counc Repub China B 25:158-65, 2001.
- Limtrakul Pn P, Anuchapreeda S, Lipigomgoson S and Dunn FW: Inhibition of carcinogen induced c-Ha-ras and c-fos proto-oncogenes expression by dietary curcumin. BMC Cancer 1: 1, 2001.
- Sindhwani P, Hampton JA, Baig MM, Keck R and Selman SH: Curcumin prevents intravesical tumor implantation of the MBT-2 tumor cell line in OH mice. J Urol 166: 1498-501, 2001.
- Lin CC, Lu YP, Lou YR, Ho CT, Newmark HH, MacDonald C, Singletary KW and Huang MT: Inhibition by dietary dibenzoylmethane of mammary gland proliferation, formation of DMBADNA adducts in mammary glands, and mammary tumorigenesis in Sencar mice. Cancer Lett 168: 125-32, 2001.
- Liao S, Lin J, Dang MT, Zhang H, Kao YH, Fukuchi J and Hiipakka RA: Growth suppression of hamster flank organs by topical application of catechins, alizarin, curcumin, and myristoleic acid. Arch Dermatol Res 293: 200-5, 2001.
- Collett GP, Robson CN, Mathers JC and Campbell FC: curcumin modifies Apc(min) apoptosis resistance and inhibits 2-amino 1-methyl-6-phenylimidazo[4,5b]pyridine (PhIP) induced tumour formation in Apc(min) mice. Carcinogenesis 22: 821-5, 2001.
- Hergenhahn M, Soto U, Weninger A, Polack A, Hsu CH, Cheng AL and Rosl F: The chemopreventive compound curcumin is an efficient inhibitor of Epstein-Barr virus BZLFl transcription in Raji DR-LUC cells. Mol Carcinog 33: 137-45, 2002.
- Inano H and Onoda M: Prevention of radiation-induced mammary tumors. Int J Radiat Oncol Biol Phys 52: 212-23, 2002.
- Li N, Chen X, Liao J, Yang G, Wang S, Josephson Y, Han C, Chen J, Huang MT and Yang CS: Inhibition of 7,12-dimethylbenz[a] anthracene (DMBA)-induced oral carcinogenesis in hamsters by tea and curcumin. Carcinogenesis 23: 1307-13, 2002.
- Huang MT, Deschner EE, Newmark HL, Wang ZY, Ferraro TA and Conney AH: Effect of dietary curcumin and ascorbyl palmitate on azoxymethanol-induced colonic epithelial cell proliferation and focal areas of dysplasia. Cancer Lett 64: 117-21, 1992.
- Inano H and Onoda M: Radioprotective action of curcumin extracted from Curcuma longa Linn: inhibitory effect on formation of urinary 8-hydroxy-2’deoxyguanosine, tumorigenesis, but not mortality, induced by gamma-ray irradiation. Int J Radiat Oncol Biol Phys 53: 735-43, 2002.
- Mahmoud NN, Carothers AM, Grunberger D, Bilinski RT, Churchill MR, Martucci C, Newmark HL and Bertagnolli MM: Plant phenolics decrease intestinal tumors in an animal model of familial adenomatous polyposis. Carcinogenesis 21: 921-7, 2000.
- Busquets S, Carbo N, Almendro V, Quiles MT, Lopez-Soriano FJ and Argiles JM: Curcumin, a natural product present in turmeric, decreases tumor growth but does not behave as an anticachectic compound in a rat model. Cancer Lett 167: 33-8, 2001.
- Dorai T, Cao YC, Dorai B, Buttyan R and Katz AE: Therapeutic potential of curcumin in human prostate cancer. III. Curcumin inhibits proliferation, induces apoptosis, and inhibits angiogenesis of LNCaP prostate cancer cells in vivo. Prostate 47: 293-303, 2001.
- Menon LG, Kuttan R and Kuttan G: Inhibition of lung metastasis in mice induced by B16F10 melanoma cells by polyphenolic compounds. Cancer Lett 95: 221-5, 1995.
- Food, Nutrition and the Prevention of Cancer: A Global Perspective. Washington DC: World Cancer Research Fund /American Institute for Cancer Research 1997.
- 337 Dyer JL, Khan SZ, Bilmen JG, Hawtin SR, Wheatley M, Javed MU and Michelangeli F: Curcumin: a new cell-permeant inhibitor of the inositol 1,4,5-trisphosphate receptor. Cell Calcium 31: 45-52, 2002.
Related Information
Videos
Links
- STUDY OF GEMCITABINE-BASED CHEMOTHERAPY PLUS CURCUMIN
- CURCUMIN ENHANCES THE CYTOTOXIC AND CHEMO-SENSITISING EFFECTS OF LENALIDOMIDE IN HUMAN MULTIPLE MYELOMA CELLS
- CONSUMPTION OF THE PUTATIVE CHEMOPREVENTIVE AGENT CURCUMIN BY CANCER PATIENTS
- PHASE I CLINICAL TRIAL OF ORAL CURCUMIN
- CURCUMIN: A NOVEL STAT 3 PATHWAY INHIBITOR FOR CHEMOPREVENTION OF LUNG CANCER
- CURCUMIN FOR RADIATION DERMATITIS
- ANTI-CANCER POTENTIAL OF CURCUMIN






